

Deficiency of protocadherin 9 leads to reduction in positive emotional behaviour
- PMID: 35831353
- PMCID: PMC9279467
- DOI: 10.1038/s41598-022-16106-5
Deficiency of protocadherin 9 leads to reduction in positive emotional behaviour
Abstract
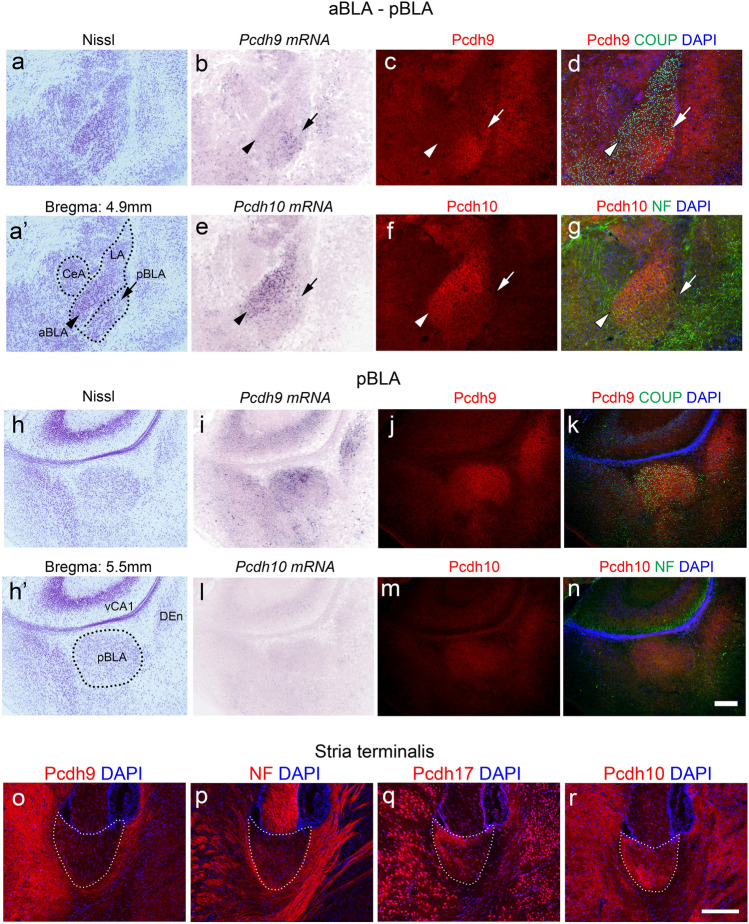
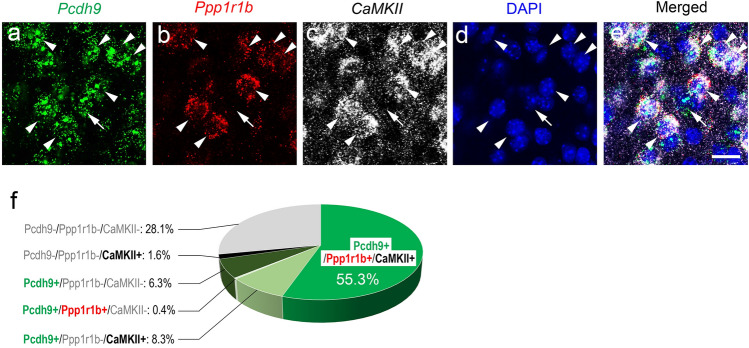
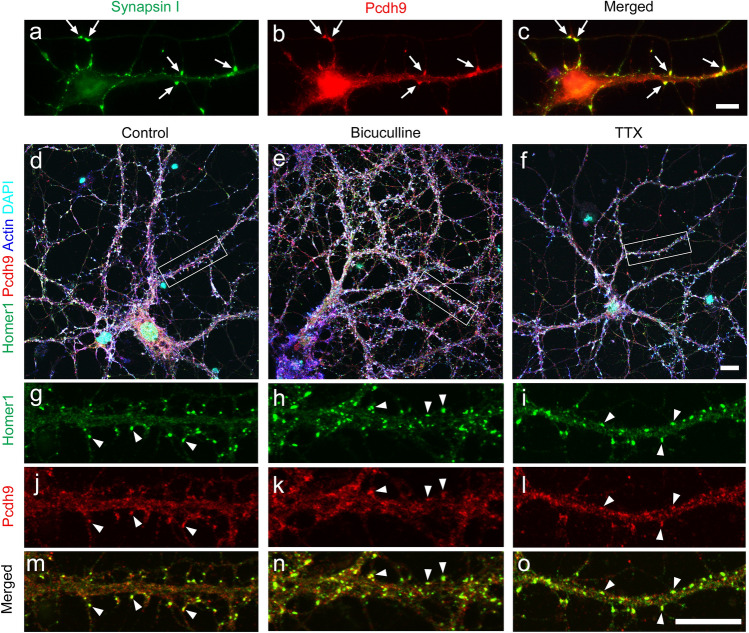
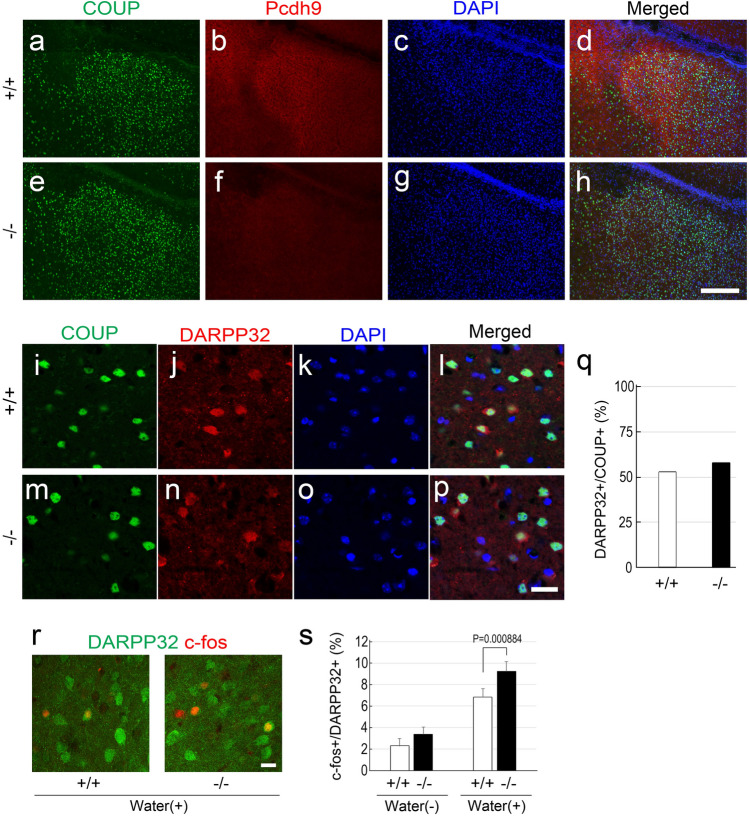
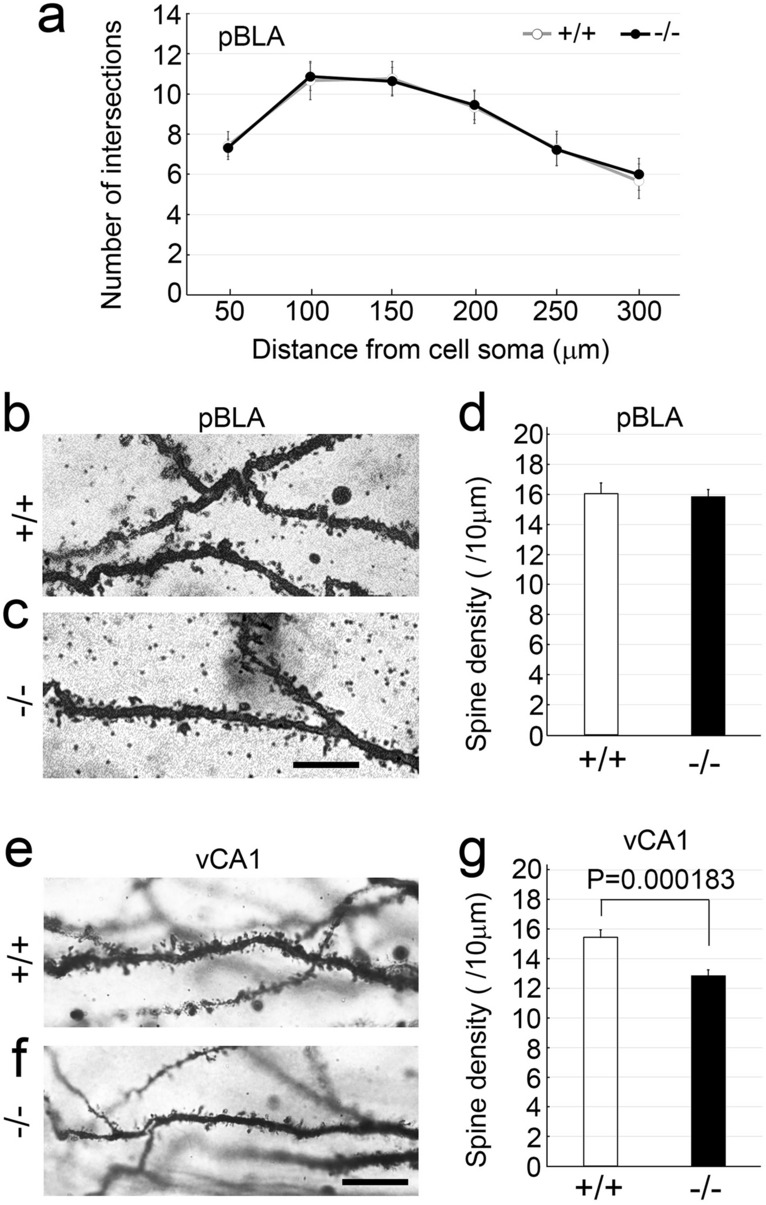
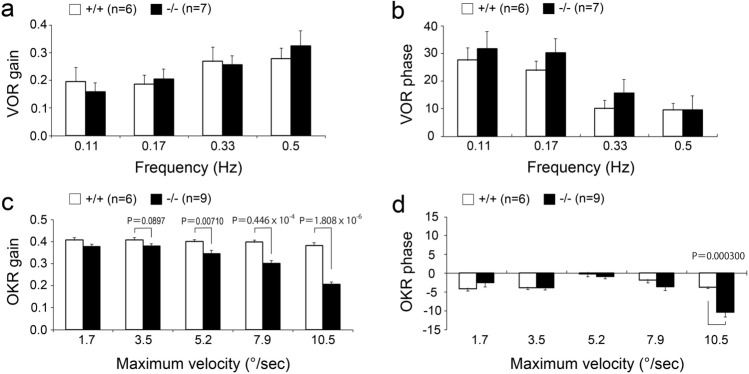
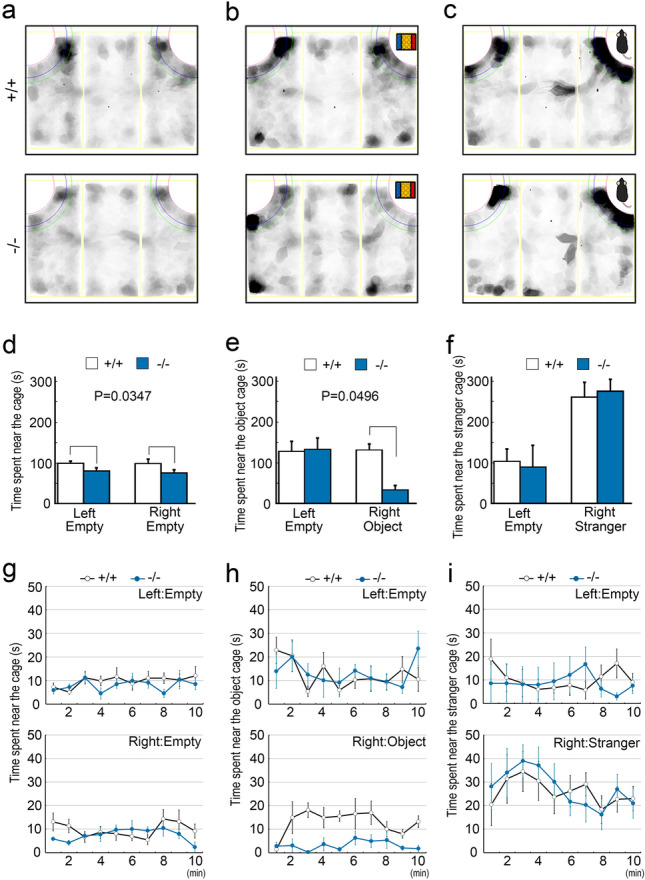
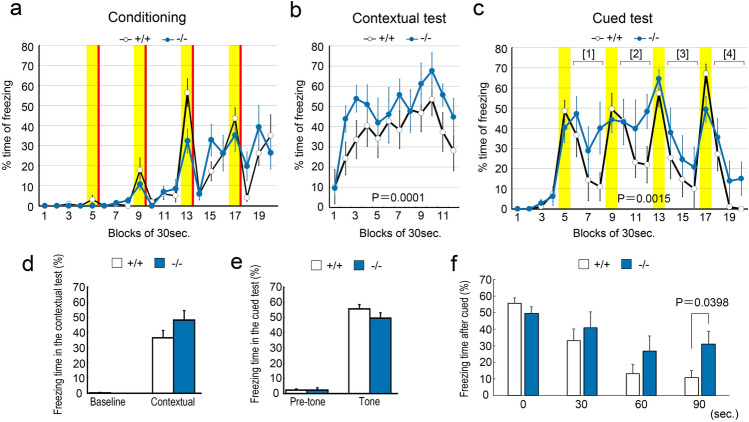
Protocadherin 9 (Pcdh9) is a member of the cadherin superfamily and is uniquely expressed in the vestibular and limbic systems; however, its physiological role remains unclear. Here, we studied the expression of Pcdh9 in the limbic system and phenotypes of Pcdh9-knock-out mice (Pcdh9 KO mice). Pcdh9 mRNA was expressed in the fear extinction neurons that express protein phosphatase 1 regulatory subunit 1 B (Ppp1r1b) in the posterior part of the basolateral amygdala (pBLA), as well as in the Cornu Ammonis (CA) and Dentate Gyrus (DG) neurons of the hippocampus. We show that the Pcdh9 protein was often localised at synapses. Phenotypic analysis of Pcdh9 KO mice revealed no apparent morphological abnormalities in the pBLA but a decrease in the spine number of CA neurons. Further, the Pcdh9 KO mice were related to features such as the abnormal optokinetic response, less approach to novel objects, and reduced fear extinction during recovery from the fear. These results suggest that Pcdh9 is involved in eliciting positive emotional behaviours, possibly via fear extinction neurons in the pBLA and/or synaptic activity in the hippocampal neurons, and normal optokinetic eye movement in brainstem optokinetic system-related neurons.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Weiner KMMJA. In: The cadherin superfamily. Suzuki ST, Hirano S, editors. Springer; 2016. pp. 195–221.
-
- Jontes JD. In: The cadherin superfamily. Suzuki ST, Hirano S, editors. Springer; 2016. pp. 223–249.
-
- Hirano S, Imai-Okano K. In: The cadherin superfamily. Suzuki ST, Hirano S, editors. Springer; 2016. pp. 399–421.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
