

Origin, Development, and Synaptogenesis of Cortical Interneurons
- PMID: 35833090
- PMCID: PMC9272671
- DOI: 10.3389/fnins.2022.929469
Origin, Development, and Synaptogenesis of Cortical Interneurons
Abstract
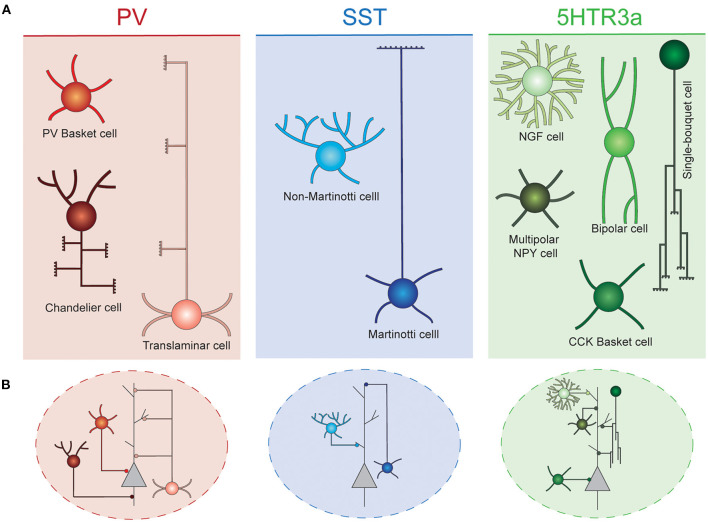
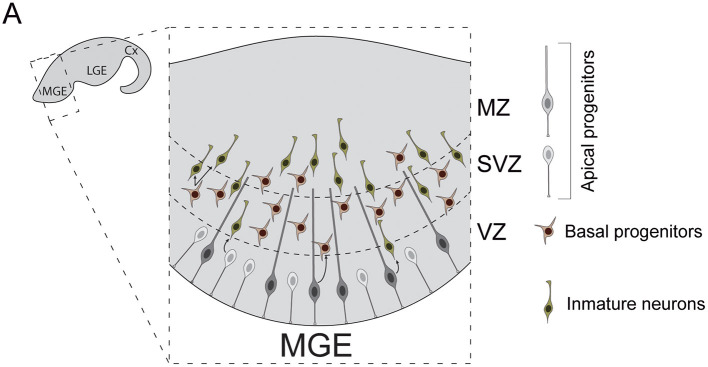
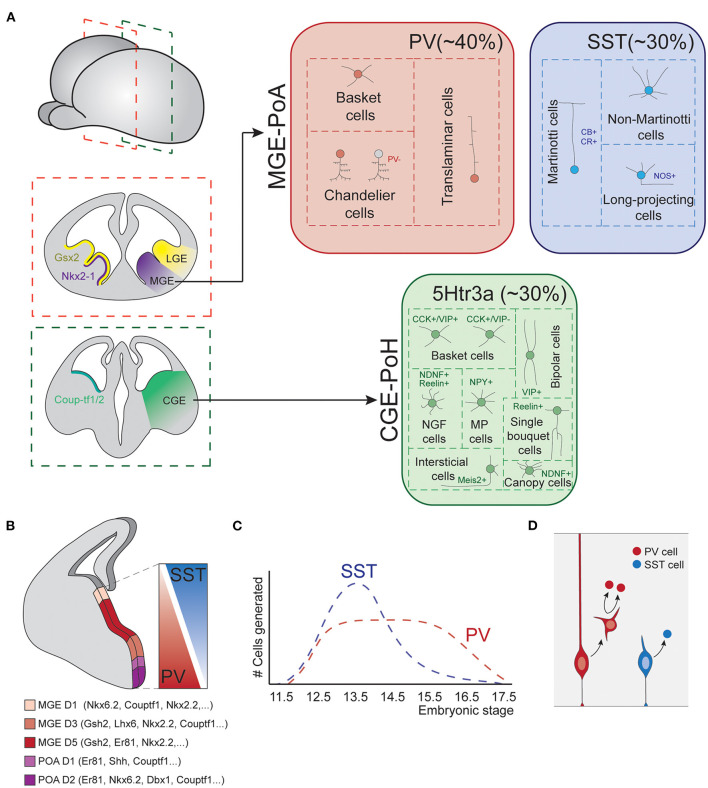
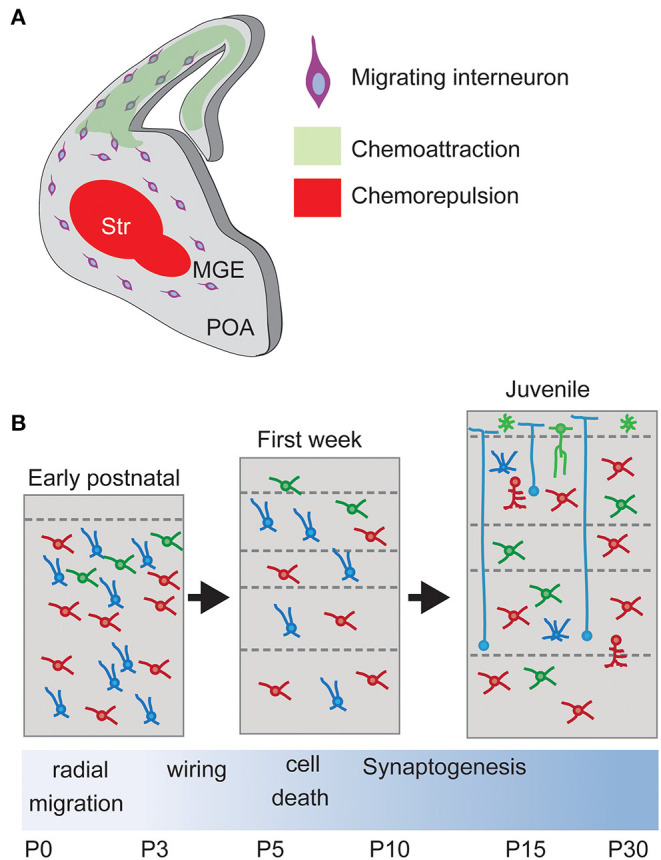
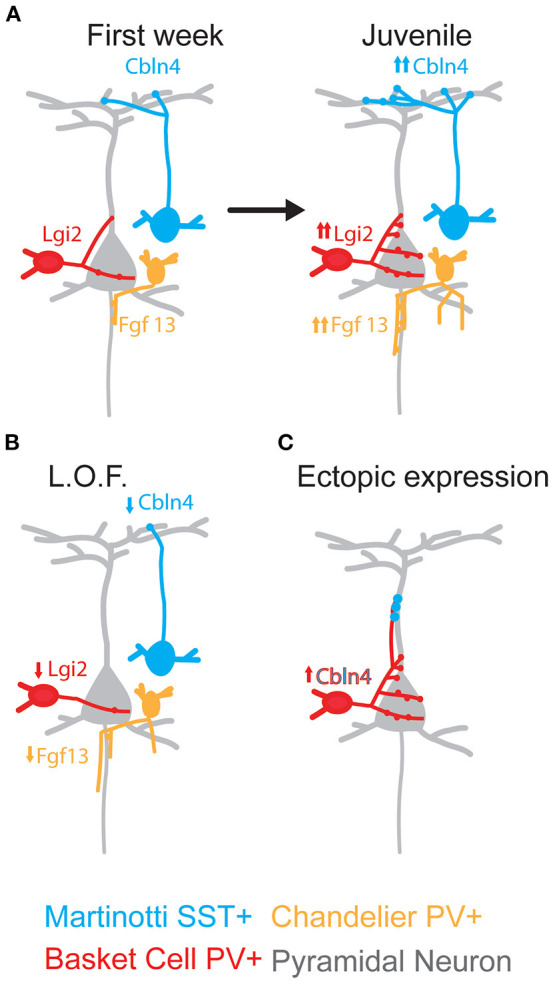
The mammalian cerebral cortex represents one of the most recent and astonishing inventions of nature, responsible of a large diversity of functions that range from sensory processing to high-order cognitive abilities, such as logical reasoning or language. Decades of dedicated study have contributed to our current understanding of this structure, both at structural and functional levels. A key feature of the neocortex is its outstanding richness in cell diversity, composed by multiple types of long-range projecting neurons and locally connecting interneurons. In this review, we will describe the great diversity of interneurons that constitute local neocortical circuits and summarize the mechanisms underlying their development and their assembly into functional networks.
Keywords: brain; cell death; cortex; development; interneuron; neuron; synaptogenesis.
Copyright © 2022 Llorca and Deogracias.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources