

BTN3A Targeting Vγ9Vδ2 T Cells Antimicrobial Activity Against Coxiella burnetii-Infected Cells
- PMID: 35833118
- PMCID: PMC9272908
- DOI: 10.3389/fimmu.2022.915244
BTN3A Targeting Vγ9Vδ2 T Cells Antimicrobial Activity Against Coxiella burnetii-Infected Cells
Abstract
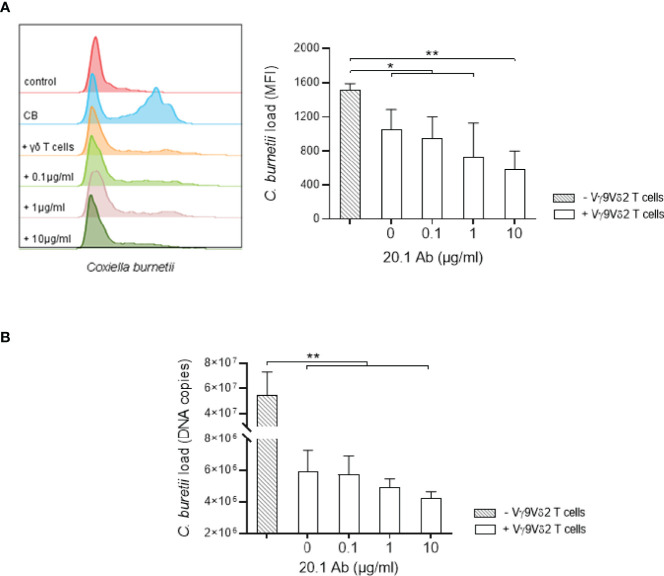
Vγ9Vδ2 T cells have been reported to participate to the immune response against infectious diseases such as the Q fever caused by Coxiella burnetii infection. Indeed, the number and proportion of Vγ9Vδ2 T cells are increased during the acute phase of Q fever. Human Vγ9Vδ2 T cell responses are triggered by phosphoantigens (pAgs) produced by pathogens and malignant cells, that are sensed via the membrane receptors butyrophilin-3A1 (BTN3A1) and -2A1 (BTN2A1). Here, by using CRISPR-Cas9 inactivation in THP-1 cells, we show that BTN3A and BTN2A are required to Vγ9Vδ2 T cell response to C. burnetii infection, though not directly involved in the infection process. Furthermore, C. burnetii-infected monocytes display increased BTN3A and BTN2A expression and induce Vγ9Vδ2 T cell activation that can be inhibited by specific antagonist mAb. More importantly, we show that the antimicrobial functions of Vγ9Vδ2 T cells towards C. burnetii are enhanced in the presence of an BTN3A activating antibody. This supports the role of Vγ9Vδ2 T cells in the control of C. burnetii infection and argues in favor of targeting these cells as an alternative treatment strategy for infectious diseases caused by intracellular bacteria.
Keywords: Coxiella burnetii; Vγ9Vδ2 T cells; antimicrobial immunity; butyrophilin; therapeutic approaches.
Copyright © 2022 Gay, Mezouar, Cano, Foucher, Gabriac, Fullana, Madakamutil, Mège and Olive.
Conflict of interest statement
DO is cofounder and shareholder of Imcheck Therapeutics, Emergence Therapeutics, Alderaan Biotechnology and Stealth IO. CC, EF, MG, MF, and LM are employees and shareholders of Imcheck Therapeutics. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.The authors declare that this study received funding from ImCheck Therapeutics. The funder had the following involvement in the study: study design, collection, analysis, interpretation of data and the writing of this article.
Figures
Similar articles
-
Role of Vγ9vδ2 T lymphocytes in infectious diseases.Front Immunol. 2022 Jul 18;13:928441. doi: 10.3389/fimmu.2022.928441. eCollection 2022. Front Immunol. 2022. PMID: 35924233 Free PMC article. Review.
-
BTN2A1, an immune checkpoint targeting Vγ9Vδ2 T cell cytotoxicity against malignant cells.Cell Rep. 2021 Jul 13;36(2):109359. doi: 10.1016/j.celrep.2021.109359. Cell Rep. 2021. PMID: 34260935
-
Butyrophilin-2A1 Directly Binds Germline-Encoded Regions of the Vγ9Vδ2 TCR and Is Essential for Phosphoantigen Sensing.Immunity. 2020 Mar 17;52(3):487-498.e6. doi: 10.1016/j.immuni.2020.02.014. Epub 2020 Mar 9. Immunity. 2020. PMID: 32155411 Free PMC article.
-
Butyrophilin 3A (BTN3A, CD277)-specific antibody 20.1 differentially activates Vγ9Vδ2 TCR clonotypes and interferes with phosphoantigen activation.Eur J Immunol. 2017 Jun;47(6):982-992. doi: 10.1002/eji.201646818. Epub 2017 Apr 27. Eur J Immunol. 2017. PMID: 28386905
-
Phosphoantigen recognition by Vγ9Vδ2 T cells.Eur J Immunol. 2024 Nov;54(11):e2451068. doi: 10.1002/eji.202451068. Epub 2024 Aug 15. Eur J Immunol. 2024. PMID: 39148158 Review.
Cited by
-
Cytometry profiling of ex vivo recall responses to Coxiella burnetii in previously naturally exposed individuals reveals long-term changes in both adaptive and innate immune cellular compartments.Front Immunol. 2023 Oct 11;14:1249581. doi: 10.3389/fimmu.2023.1249581. eCollection 2023. Front Immunol. 2023. PMID: 37885896 Free PMC article.
-
Pathogenicity and Virulence of Coxiella burnetii: Focus on Q fever.Virulence. 2025 Dec;16(1):2495842. doi: 10.1080/21505594.2025.2495842. Epub 2025 Jun 19. Virulence. 2025. PMID: 40536474 Free PMC article. Review.
-
Role of Vγ9vδ2 T lymphocytes in infectious diseases.Front Immunol. 2022 Jul 18;13:928441. doi: 10.3389/fimmu.2022.928441. eCollection 2022. Front Immunol. 2022. PMID: 35924233 Free PMC article. Review.
-
Reducing farnesyl diphosphate synthase levels activates Vγ9Vδ2 T cells and improves tumor suppression in murine xenograft cancer models.Front Immunol. 2022 Oct 5;13:1012051. doi: 10.3389/fimmu.2022.1012051. eCollection 2022. Front Immunol. 2022. PMID: 36275712 Free PMC article.
-
Macrophages and γδ T cells interplay during SARS-CoV-2 variants infection.Front Immunol. 2022 Dec 19;13:1078741. doi: 10.3389/fimmu.2022.1078741. eCollection 2022. Front Immunol. 2022. PMID: 36601113 Free PMC article.
References
-
- Morita CT, Jin C, Sarikonda G, Wang H. Nonpeptide Antigens, Presentation Mechanisms, and Immunological Memory of Human Vγ2vδ2 T Cells: Discriminating Friend From Foe Through the Recognition of Prenyl Pyrophosphate Antigens. Immunol Rev (2007) 215:59–76. doi: 10.1111/j.1600-065X.2006.00479.x - DOI - PubMed
-
- Balbi B, Valle MT, Oddera S, Giunti O, Manca F, Rossi GA, et al. . T-Lymphocytes With γδ+ Vδ2+ Antigen Receptors are Present in Increased Proportions in a Fraction of Patients With Tuberculosis or With Sarcoidosis. Am Rev Respir Dis (1993) 148:1685–90. doi: 10.1164/ajrccm/148.6_Pt_1.1685 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases