

Pathogenic autoantibodies to IFN-γ act through the impedance of receptor assembly and Fc-mediated response
- PMID: 35833912
- PMCID: PMC9287643
- DOI: 10.1084/jem.20212126
Pathogenic autoantibodies to IFN-γ act through the impedance of receptor assembly and Fc-mediated response
Abstract
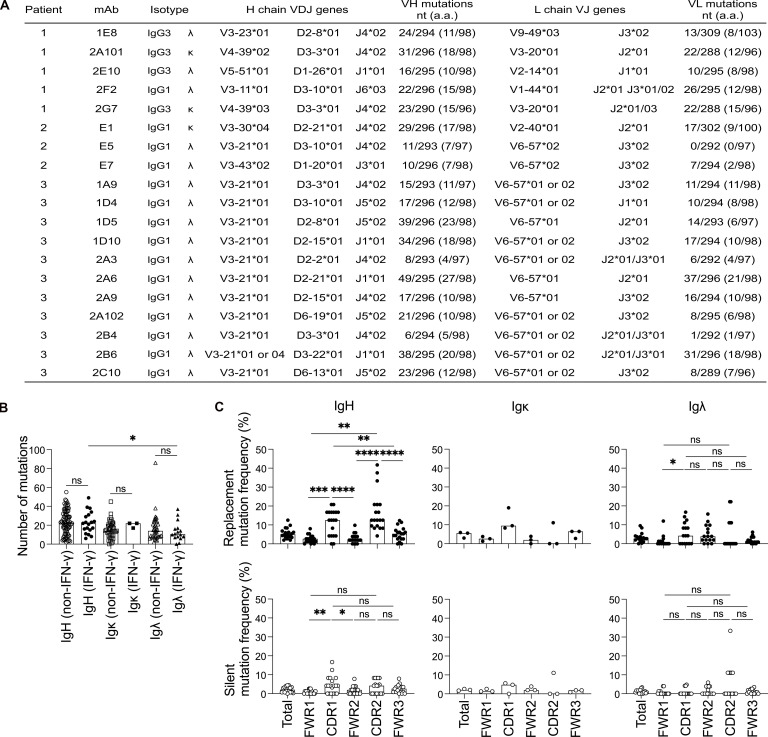
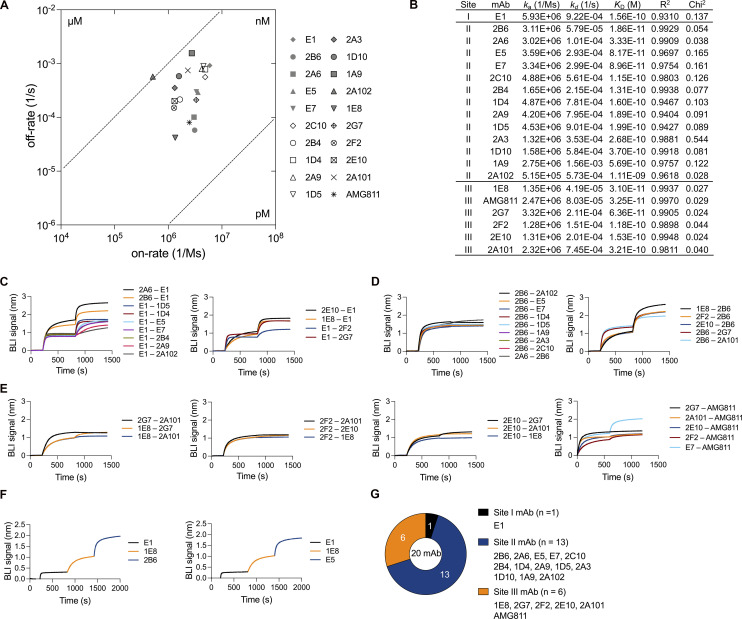
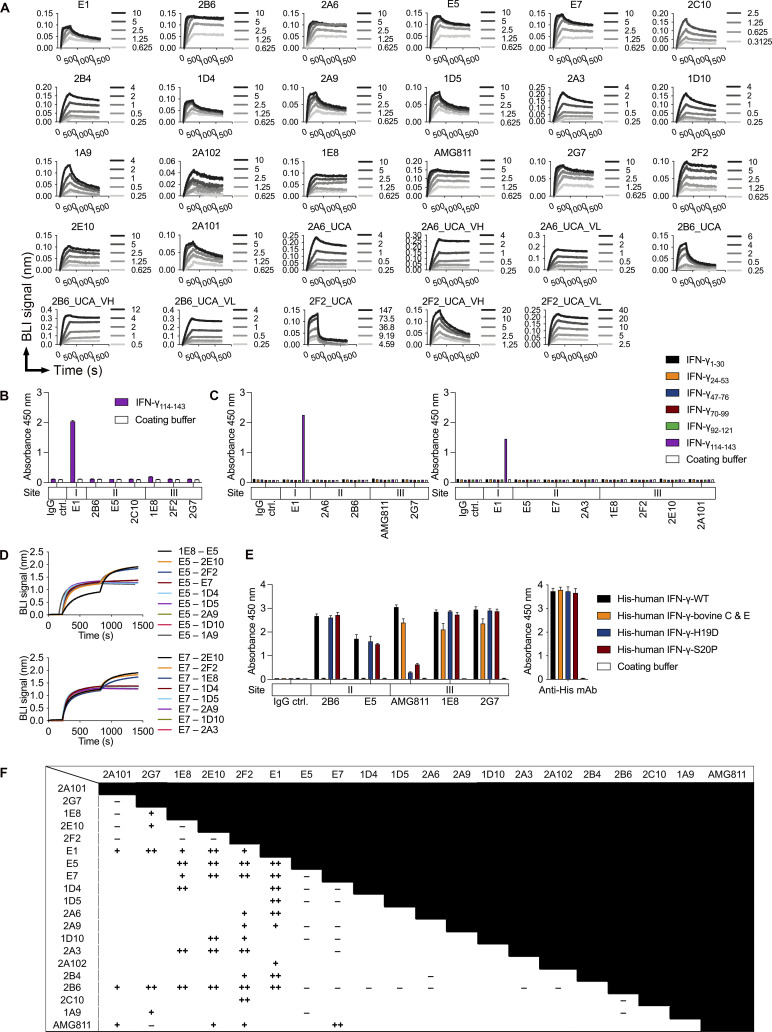
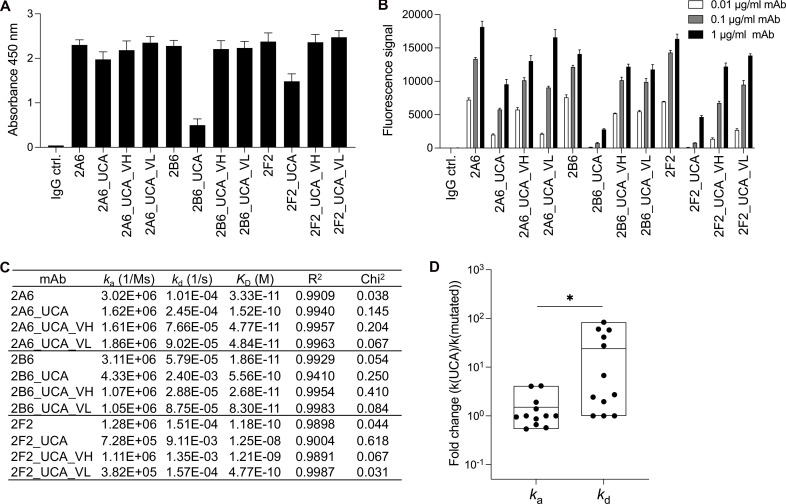
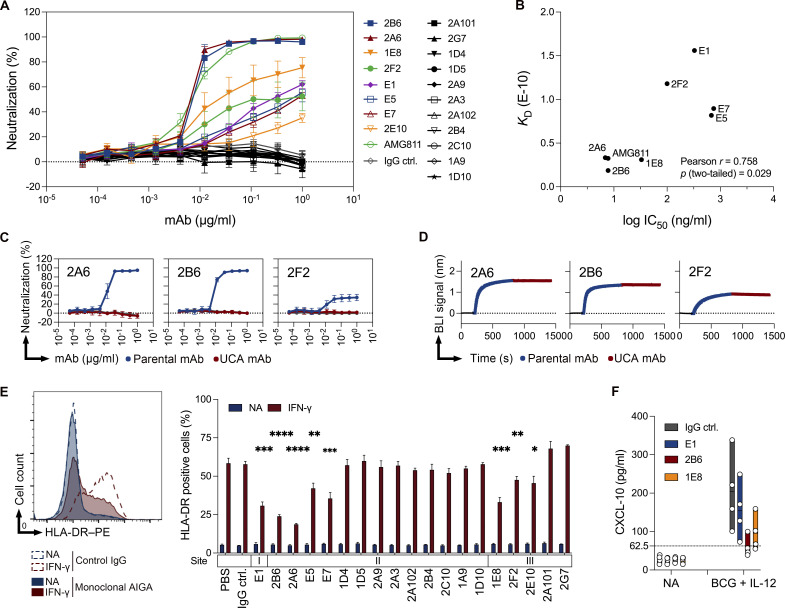
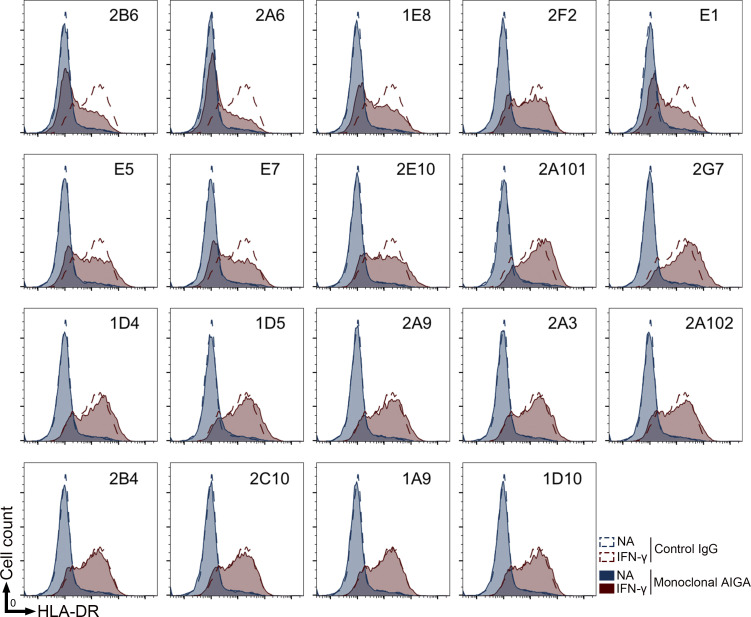
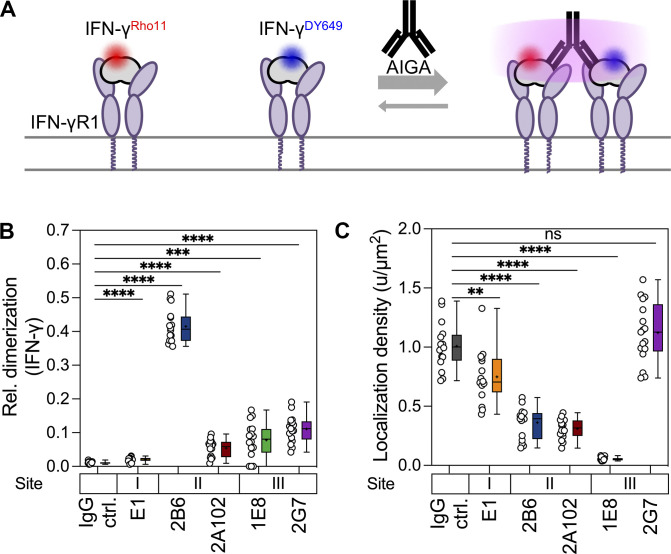
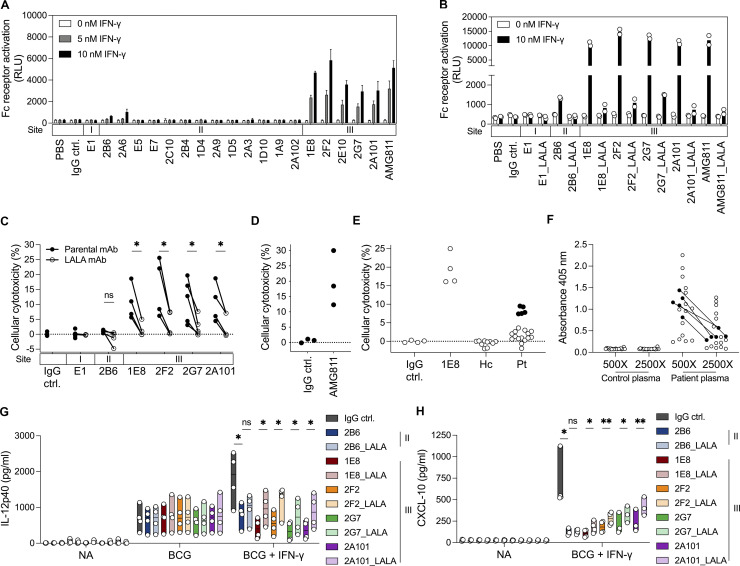
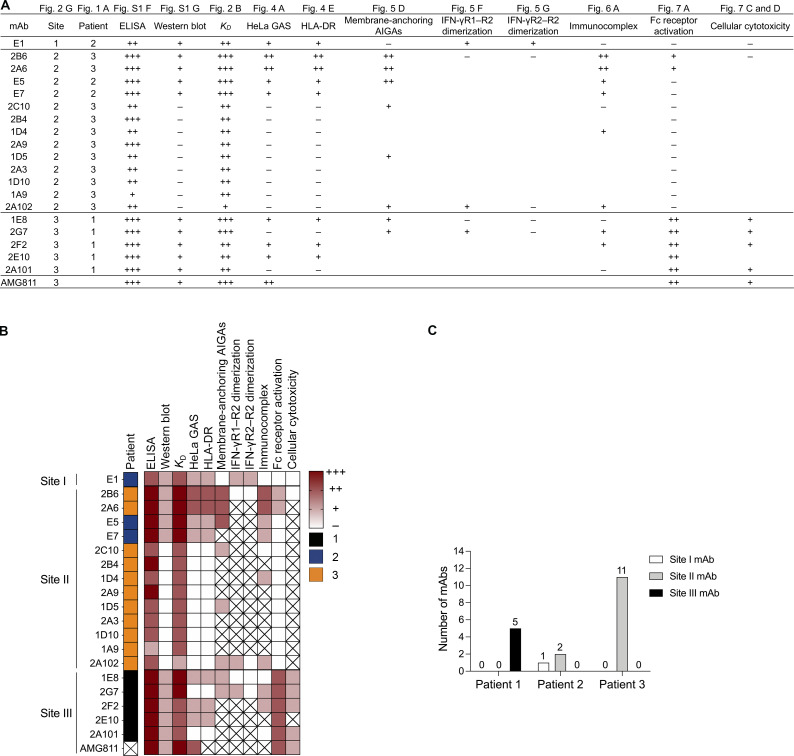
Anti-interferon (IFN)-γ autoantibodies (AIGAs) are a pathogenic factor in late-onset immunodeficiency with disseminated mycobacterial and other opportunistic infections. AIGAs block IFN-γ function, but their effects on IFN-γ signaling are unknown. Using a single-cell capture method, we isolated 19 IFN-γ-reactive monoclonal antibodies (mAbs) from patients with AIGAs. All displayed high-affinity (KD < 10-9 M) binding to IFN-γ, but only eight neutralized IFN-γ-STAT1 signaling and HLA-DR expression. Signal blockade and binding affinity were correlated and attributed to somatic hypermutations. Cross-competition assays identified three nonoverlapping binding sites (I-III) for AIGAs on IFN-γ. We found that site I mAb neutralized IFN-γ by blocking its binding to IFN-γR1. Site II and III mAbs bound the receptor-bound IFN-γ on the cell surface, abolishing IFN-γR1-IFN-γR2 heterodimerization and preventing downstream signaling. Site III mAbs mediated antibody-dependent cellular cytotoxicity, probably through antibody-IFN-γ complexes on cells. Pathogenic AIGAs underlie mycobacterial infections by the dual blockade of IFN-γ signaling and by eliminating IFN-γ-responsive cells.
© 2022 Shih et al.
Conflict of interest statement
Disclosures: H.-P. Shih, J.-Y. Ding, C.-H. Lin, J.-Y. Huang, and C.-L. Ku reported a patent to US 10,968,274 B2 issued. No other disclosures were reported.
Figures



Similar articles
-
Patient iPSC-Derived Macrophages to Study Inborn Errors of the IFN-γ Responsive Pathway.Cells. 2020 Feb 19;9(2):483. doi: 10.3390/cells9020483. Cells. 2020. PMID: 32093117 Free PMC article.
-
Partial interferon-gamma receptor deficiency and non-tuberculous mycobacterial lung disease.Tuberculosis (Edinb). 2006 Sep;86(5):382-5. doi: 10.1016/j.tube.2005.11.002. Epub 2006 May 8. Tuberculosis (Edinb). 2006. PMID: 16682253
-
IFN-γR1 defects: Mutation update and description of the IFNGR1 variation database.Hum Mutat. 2017 Oct;38(10):1286-1296. doi: 10.1002/humu.23302. Epub 2017 Aug 3. Hum Mutat. 2017. PMID: 28744922 Review.
-
Enhanced osteoclastogenesis in patients with MSMD due to impaired response to IFN-γ.J Allergy Clin Immunol. 2022 Jan;149(1):252-261.e6. doi: 10.1016/j.jaci.2021.05.018. Epub 2021 Jun 24. J Allergy Clin Immunol. 2022. PMID: 34176646
-
Intact IFN-γR1 expression and function distinguishes Langerhans cell histiocytosis from mendelian susceptibility to mycobacterial disease.J Clin Immunol. 2014 Jan;34(1):84-93. doi: 10.1007/s10875-013-9959-1. Epub 2013 Nov 20. J Clin Immunol. 2014. PMID: 24254535 Review.
Cited by
-
A sensitive assay for measuring whole-blood responses to type I IFNs.Proc Natl Acad Sci U S A. 2024 Oct;121(40):e2402983121. doi: 10.1073/pnas.2402983121. Epub 2024 Sep 23. Proc Natl Acad Sci U S A. 2024. PMID: 39312669 Free PMC article.
-
Human immunity to fungal infections.J Exp Med. 2025 Jun 2;222(6):e20241215. doi: 10.1084/jem.20241215. Epub 2025 Apr 15. J Exp Med. 2025. PMID: 40232283 Review.
-
A Proteomics Outlook on the Molecular Effectors of CAR-T Cell Therapy in Cancer Management.J Proteome Res. 2025 Jun 6;24(6):2571-2583. doi: 10.1021/acs.jproteome.4c00930. Epub 2025 Mar 6. J Proteome Res. 2025. PMID: 40047620 Free PMC article. Review.
-
A homogeneous bioluminescent inhibition immunoassay to detect anti-interferon gamma antibodies.Clin Exp Immunol. 2024 Nov 12;218(3):283-290. doi: 10.1093/cei/uxae055. Clin Exp Immunol. 2024. PMID: 38975719 Free PMC article.
-
Autoantibodies to Erythropoietin Receptor and Clinical Outcomes in Patients With Type 2 Diabetes and CKD: A Post Hoc Analysis of CREDENCE Trial.Kidney Int Rep. 2023 Dec 1;9(2):347-355. doi: 10.1016/j.ekir.2023.11.024. eCollection 2024 Feb. Kidney Int Rep. 2023. PMID: 38344715 Free PMC article.
References
-
- Aoki, A., Sakagami T., Yoshizawa K., Shima K., Toyama M., Tanabe Y., Moro H., Aoki N., Watanabe S., Koya T., et al. . 2018. Clinical significance of interferon-γ neutralizing autoantibodies against disseminated nontuberculous mycobacterial disease. Clin. Infect. Dis. 66:1239–1245. 10.1093/cid/cix996 - DOI - PubMed
-
- Bastard, P., Orlova E., Sozaeva L., Levy R., James A., Schmitt M.M., Ochoa S., Kareva M., Rodina Y., Gervais A., et al. . 2021. Preexisting autoantibodies to type I IFNs underlie critical COVID-19 pneumonia in patients with APS-1. J. Exp. Med. 218:e20210554. 10.1084/jem.20210554 - DOI - PMC - PubMed
-
- Boedigheimer, M.J., Martin D.A., Amoura Z., Sanchez-Guerrero J., Romero-Diaz J., Kivitz A., Aranow C., Chan T.M., Chong Y.B., Chiu K., et al. . 2017. Safety, pharmacokinetics and pharmacodynamics of AMG 811, an anti-interferon-γ monoclonal antibody, in SLE subjects without or with lupus nephritis. Lupus Sci. Med. 4:e000226. 10.1136/lupus-2017-000226 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
