

A taxonomically representative strain collection to explore xenobiotic and secondary metabolism in bacteria
- PMID: 35834592
- PMCID: PMC9282458
- DOI: 10.1371/journal.pone.0271125
A taxonomically representative strain collection to explore xenobiotic and secondary metabolism in bacteria
Abstract
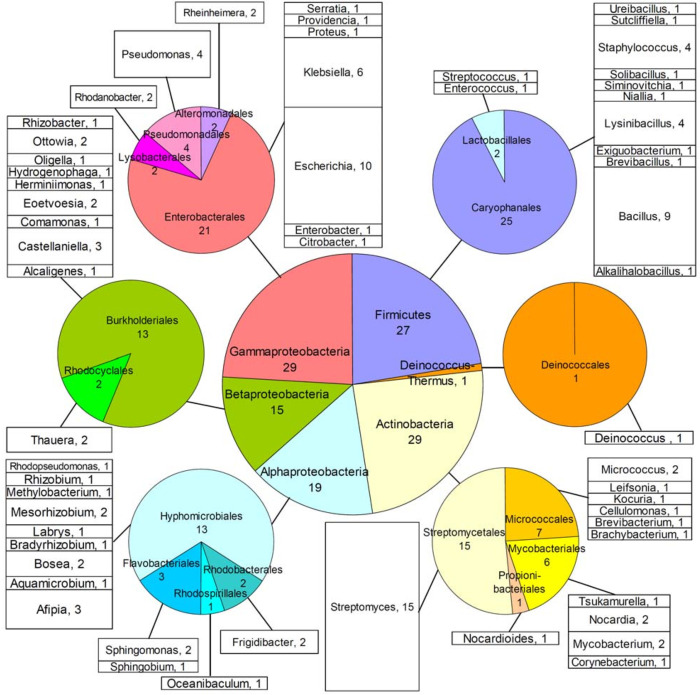
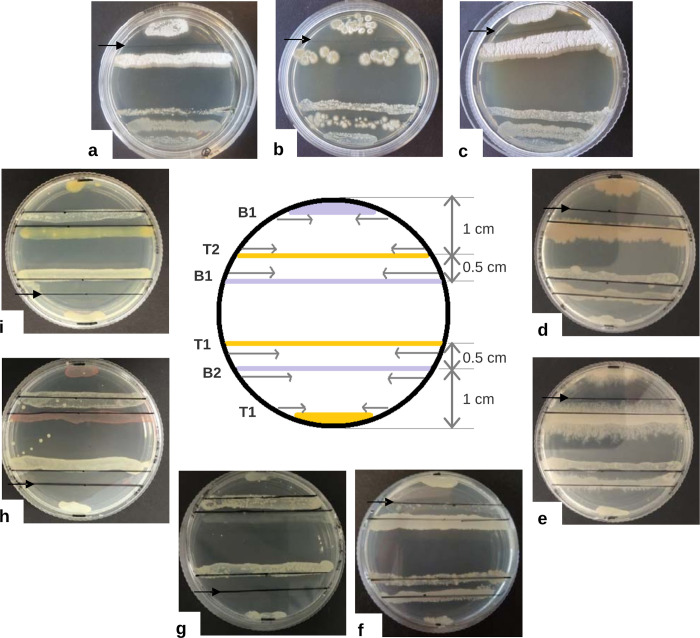
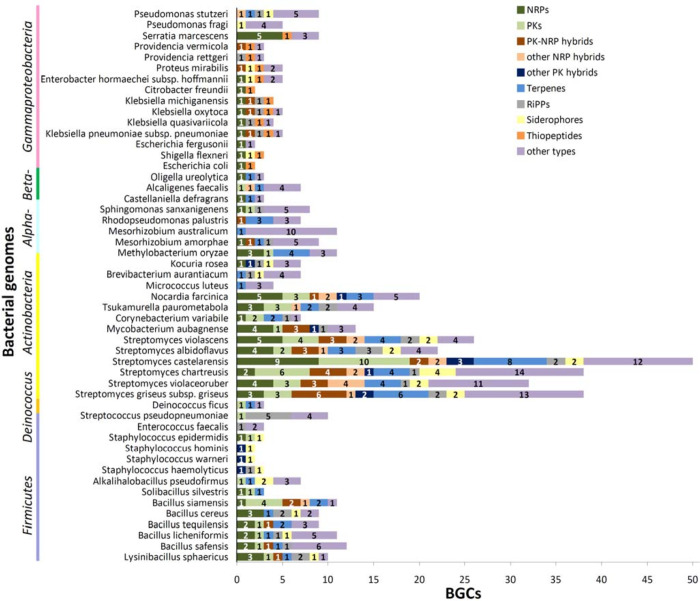
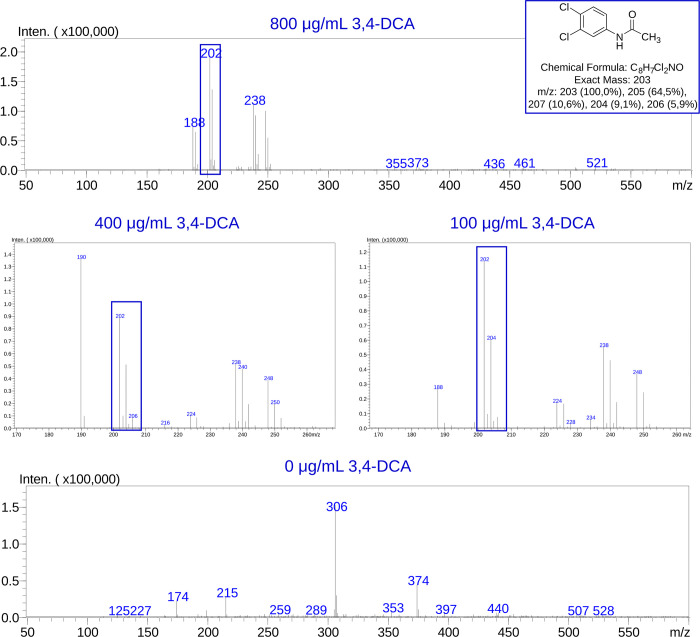
Bacteria employ secondary metabolism to combat competitors, and xenobiotic metabolism to survive their chemical environment. This project has aimed to introduce a bacterial collection enabling comprehensive comparative investigations of those functions. The collection comprises 120 strains (Proteobacteria, Actinobacteria and Firmicutes), and was compiled on the basis of the broad taxonomic range of isolates and their postulated biosynthetic and/or xenobiotic detoxification capabilities. The utility of the collection was demonstrated in two ways: first, by performing 5144 co-cultures, recording inhibition between isolates and employing bioinformatics to predict biosynthetic gene clusters in sequenced genomes of species; second, by screening for xenobiotic sensitivity of isolates against 2-benzoxazolinone and 2-aminophenol. The co-culture medium of Bacillus siamensis D9 and Lysinibacillus sphaericus DSM 28T was further analysed for possible antimicrobial compounds, using liquid chromatography-mass spectrometry (LC-MS), and guided by computational predictions and the literature. Finally, LC-MS analysis demonstrated N-acetylation of 3,4-dichloroaniline (a toxic pesticide residue of concern) by the actinobacterium Tsukamurella paurometabola DSM 20162T which is highly tolerant of the xenobiotic. Microbial collections enable "pipeline" comparative screening of strains: on the one hand, bacterial co-culture is a promising approach for antibiotic discovery; on the other hand, bioremediation is effective in combating pollution, but requires knowledge of microbial xenobiotic metabolism. The presented outcomes are anticipated to pave the way for studies that may identify bacterial strains and/or metabolites of merit in biotechnological applications.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
