

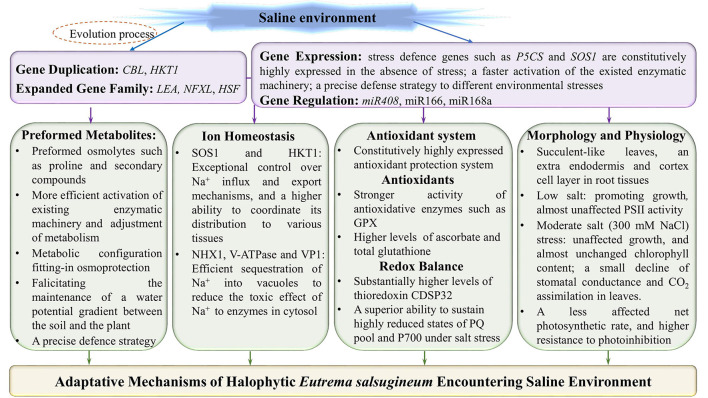
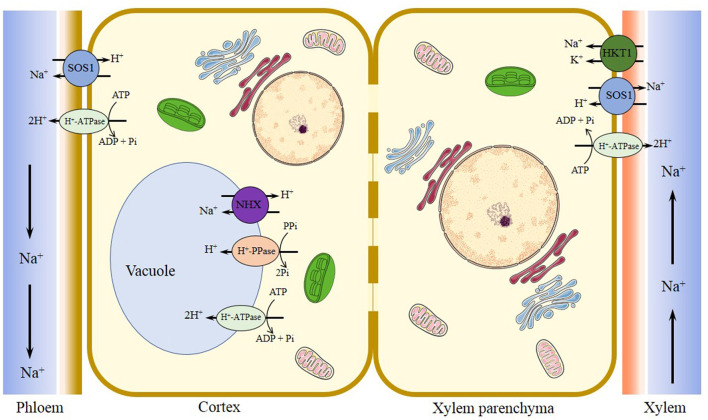
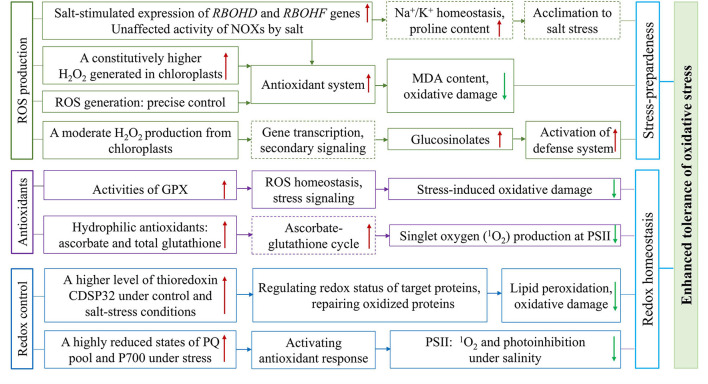
Adaptative Mechanisms of Halophytic Eutrema salsugineum Encountering Saline Environment
- PMID: 35837468
- PMCID: PMC9274170
- DOI: 10.3389/fpls.2022.909527
Adaptative Mechanisms of Halophytic Eutrema salsugineum Encountering Saline Environment
Abstract
Salt cress (Eutrema salsugineum), an Arabidopsis-related halophyte, can naturally adapt to various harsh climates and soil conditions; thus, it is considered a desirable model plant for deciphering mechanisms of salt and other abiotic stresses. Accumulating evidence has revealed that compared with Arabidopsis, salt cress possesses stomata that close more tightly and more succulent leaves during extreme salt stress, a noticeably higher level of proline, inositols, sugars, and organic acids, as well as stress-associated transcripts in unstressed plants, and they are induced rapidly under stress. In this review, we systematically summarize the research on the morphology, physiology, genome, gene expression and regulation, and protein and metabolite profile of salt cress under salt stress. We emphasize the latest advances in research on the genome adaptive evolution encountering saline environments, and epigenetic regulation, and discuss the mechanisms underlying salt tolerance in salt cress. Finally, we discuss the existing questions and opportunities for future research in halophytic Eutrema. Together, the review fosters a better understanding of the mechanism of plant salt tolerance and provides a reference for the research and utilization of Eutrema as a model extremophile in the future. Furthermore, the prospects for salt cress applied to explore the mechanism of salt tolerance provide a theoretical basis to develop new strategies for agricultural biotechnology.
Keywords: Arabidopsis relative model system; antioxidant system; gene expression; ion homeostasis; osmo-adaptation; saline adaptation; salt cress; salt stress tolerance.
Copyright © 2022 Li, Duan, Zhang, Zhao, Meng, Zhao and Zhang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
Salt cress. A halophyte and cryophyte Arabidopsis relative model system and its applicability to molecular genetic analyses of growth and development of extremophiles.Plant Physiol. 2004 Jul;135(3):1718-37. doi: 10.1104/pp.104.041723. Epub 2004 Jul 9. Plant Physiol. 2004. PMID: 15247369 Free PMC article.
-
The Reference Genome of the Halophytic Plant Eutrema salsugineum.Front Plant Sci. 2013 Mar 21;4:46. doi: 10.3389/fpls.2013.00046. eCollection 2013. Front Plant Sci. 2013. PMID: 23518688 Free PMC article.
-
Anastatica hierochuntica, an Arabidopsis Desert Relative, Is Tolerant to Multiple Abiotic Stresses and Exhibits Species-Specific and Common Stress Tolerance Strategies with Its Halophytic Relative, Eutrema (Thellungiella) salsugineum.Front Plant Sci. 2017 Jan 17;7:1992. doi: 10.3389/fpls.2016.01992. eCollection 2016. Front Plant Sci. 2017. PMID: 28144244 Free PMC article.
-
Halophytism: What Have We Learnt From Arabidopsis thaliana Relative Model Systems?Plant Physiol. 2018 Nov;178(3):972-988. doi: 10.1104/pp.18.00863. Epub 2018 Sep 20. Plant Physiol. 2018. PMID: 30237204 Free PMC article. Review.
-
Lipid remodelling: Unravelling the response to cold stress in Arabidopsis and its extremophile relative Eutrema salsugineum.Plant Sci. 2017 Oct;263:194-200. doi: 10.1016/j.plantsci.2017.07.017. Epub 2017 Jul 27. Plant Sci. 2017. PMID: 28818375 Free PMC article. Review.
Cited by
-
Physiological and transcriptomic analysis uncovers salinity stress mechanisms in a facultative crassulacean acid metabolism plant Dendrobium officinale.Front Plant Sci. 2022 Oct 6;13:1028245. doi: 10.3389/fpls.2022.1028245. eCollection 2022. Front Plant Sci. 2022. PMID: 36275597 Free PMC article.
-
Insights into the effects of saline forage on the meat quality of Tibetan sheep by metabolome and multivariate analysis.Food Chem X. 2024 Apr 26;22:101411. doi: 10.1016/j.fochx.2024.101411. eCollection 2024 Jun 30. Food Chem X. 2024. PMID: 38756473 Free PMC article.
-
Calligonum caput-medusae seedlings adapt to drought stress through changing chlorophyll fluorescence parameters.Front Plant Sci. 2025 Aug 7;16:1640412. doi: 10.3389/fpls.2025.1640412. eCollection 2025. Front Plant Sci. 2025. PMID: 40851830 Free PMC article.
References
-
- Achard P., Gong F., Cheminant S., Alioua M., Hedden P., Genschik P. (2008). The cold-inducible CBF1 factor-dependent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism. Plant Cell 20, 2117–2129. 10.1105/tpc.108.058941 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources