

A cAMP Sensor Based on Ligand-Dependent Protein Stabilization
- PMID: 35839076
- PMCID: PMC9396618
- DOI: 10.1021/acschembio.2c00333
A cAMP Sensor Based on Ligand-Dependent Protein Stabilization
Erratum in
-
Correction to "A cAMP Sensor Based on Ligand-Dependent Protein Stabilization".ACS Chem Biol. 2024 Apr 19;19(4):1022. doi: 10.1021/acschembio.4c00126. Epub 2024 Mar 8. ACS Chem Biol. 2024. PMID: 38456802 Free PMC article. No abstract available.
Abstract
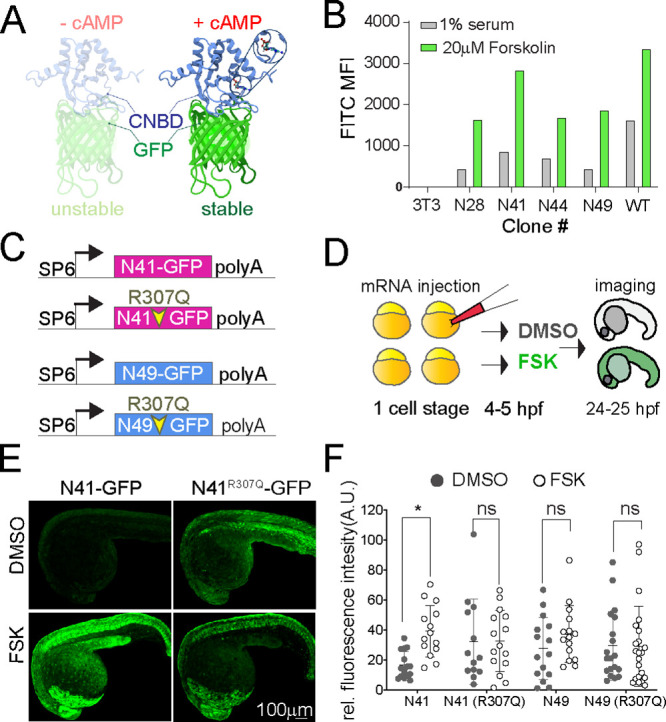
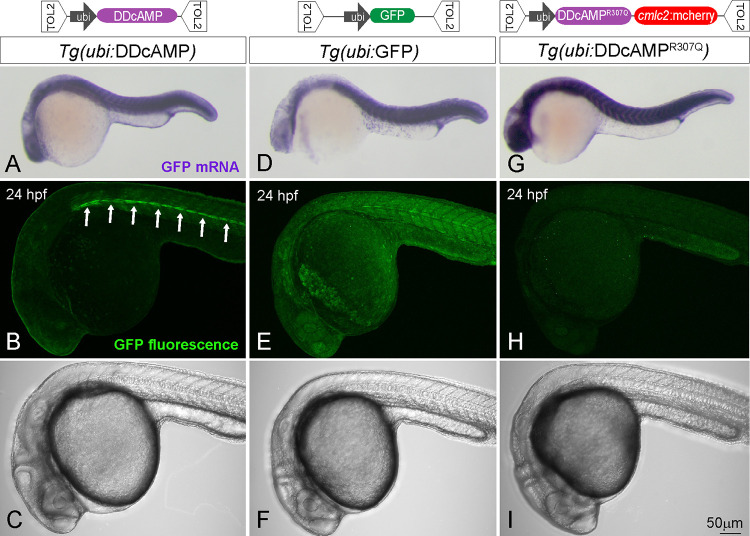
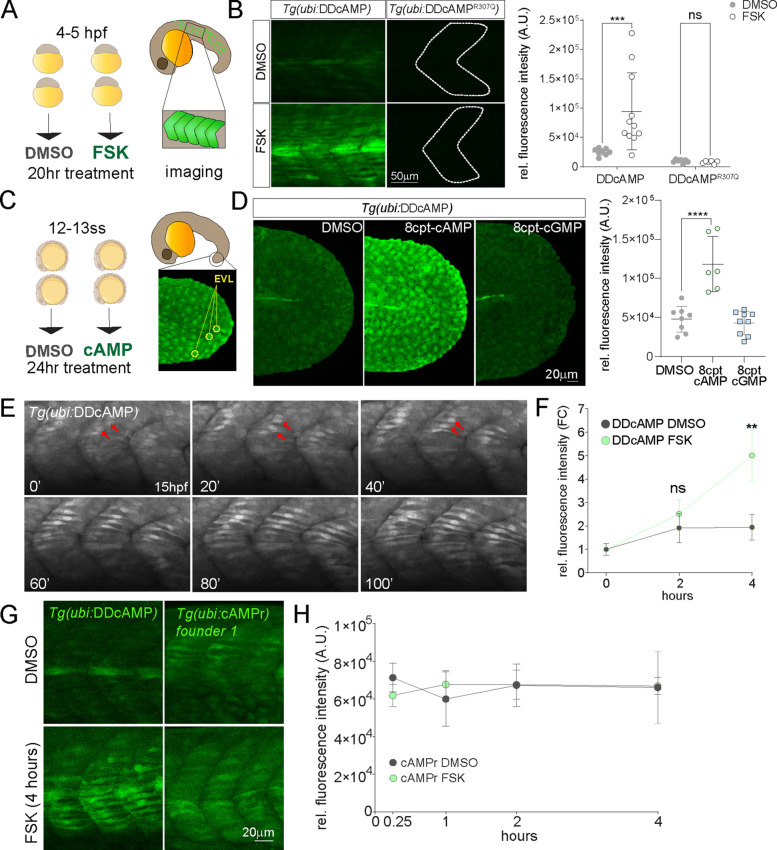
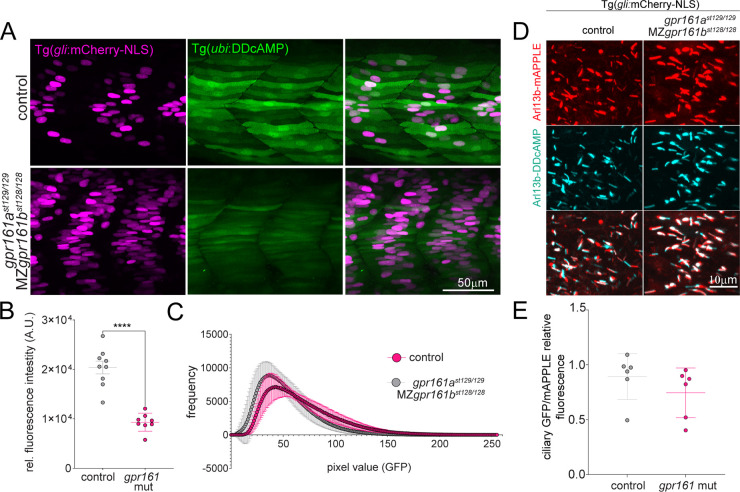
cAMP is a ubiquitous second messenger with many functions in diverse organisms. Current cAMP sensors, including Föster resonance energy transfer (FRET)-based and single-wavelength-based sensors, allow for real time visualization of this small molecule in cultured cells and in some cases in vivo. Nonetheless the observation of cAMP in living animals is still difficult, typically requiring specialized microscopes and ex vivo tissue processing. Here we used ligand-dependent protein stabilization to create a new cAMP sensor. This sensor allows specific and sensitive detection of cAMP in living zebrafish embryos, which may enable new understanding of the functions of cAMP in living vertebrates.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Morozov A.; Muzzio I. A.; Bourtchouladze R.; Van-Strien N.; Lapidus K.; Yin D.; Winder D. G.; Adams J. P.; Sweatt J. D.; Kandel E. R. Rap1 Couples CAMP Signaling to a Distinct Pool of P42/44MAPK Regulating Excitability, Synaptic Plasticity, Learning, and Memory. Neuron 2003, 39 (2), 309–325. 10.1016/S0896-6273(03)00404-5. - DOI - PubMed
-
- Surdo N. C.; Berrera M.; Koschinski A.; Brescia M.; Machado M. R.; Carr C.; Wright P.; Gorelik J.; Morotti S.; Grandi E. M.; et al. FRET Biosensor Uncovers CAMP Nano-Domains at β-Adrenergic Targets That Dictate Precise Tuning of Cardiac Contractility. Nat. Commun. 2017, 8 (1), 15031. 10.1038/ncomms15031. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
