

The potential role of HIV-1 latency in promoting neuroinflammation and HIV-1-associated neurocognitive disorder
- PMID: 35840529
- PMCID: PMC9339484
- DOI: 10.1016/j.it.2022.06.003
The potential role of HIV-1 latency in promoting neuroinflammation and HIV-1-associated neurocognitive disorder
Abstract
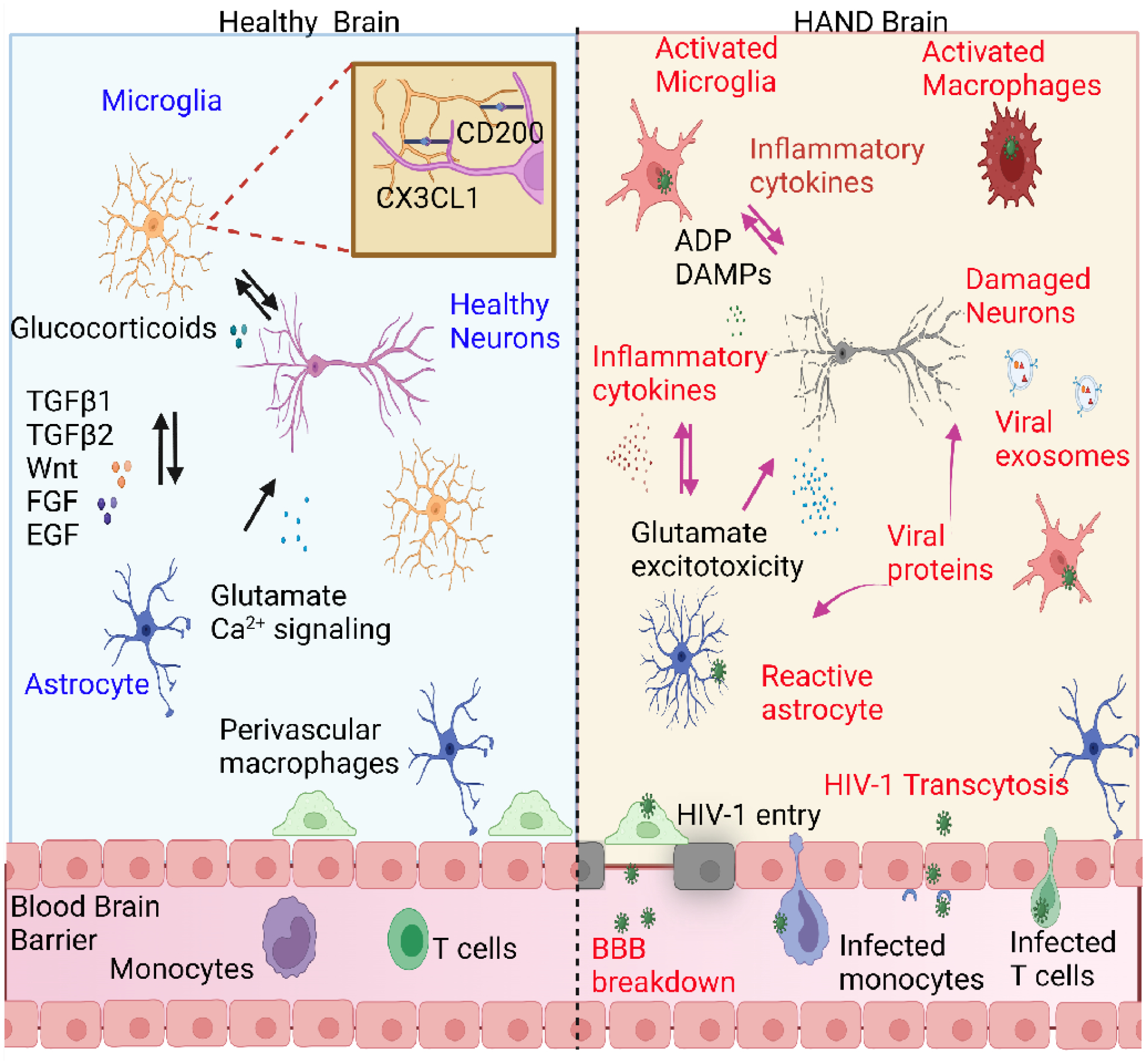
Despite potent suppression of HIV-1 viral replication in the central nervous system (CNS) by antiretroviral therapy (ART), between 15% and 60% of HIV-1-infected patients receiving ART exhibit neuroinflammation and symptoms of HIV-1-associated neurocognitive disorder (HAND) - a significant unmet challenge. We propose that the emergence of HIV-1 from latency in microglia underlies both neuroinflammation in the CNS and the progression of HAND. Recent molecular studies of cellular silencing mechanisms of HIV-1 in microglia show that HIV-1 latency can be reversed both by proinflammatory cytokines and by signals from damaged neurons, potentially creating intermittent cycles of HIV-1 reactivation and silencing in the brain. We posit that anti-inflammatory agents that also block HIV-1 reactivation, such as nuclear receptor agonists, might provide new putative therapeutic avenues for the treatment of HAND.
Keywords: HIV-1 latency; HIV-1-associated neurocognitive disorder; anti-inflammatory strategies; antiretroviral therapy; microglial cell activation.
Copyright © 2022 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Declaration of interests No interests are declared.
Figures

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
