

Expansion of mouse castration-resistant intermediate prostate stem cells in vitro
- PMID: 35841025
- PMCID: PMC9284701
- DOI: 10.1186/s13287-022-02978-x
Expansion of mouse castration-resistant intermediate prostate stem cells in vitro
Erratum in
-
Correction: Expansion of mouse castration-resistant intermediate prostate stem cells in vitro.Stem Cell Res Ther. 2022 Nov 25;13(1):522. doi: 10.1186/s13287-022-03207-1. Stem Cell Res Ther. 2022. PMID: 36434710 Free PMC article. No abstract available.
Abstract
Background: Most castration-resistant prostate cancers (CRPCs) have a luminal phenotype with high androgen receptor (AR) and prostate-specific antigen (PSA) expression. Currently, it is difficult to culture castration-resistant luminal cells with AR and PSA expression.
Methods: We formulated a custom-made medium and isolated primary cells from the prostate of adult wild-type (WT) and TRAMP mice. The cells were characterized by immunofluorescence staining, transcriptomic analysis, and qRT-PCR verification. Their self-renewal and differentiation potential in vitro and in vivo were examined. We treated the cells with androgen deprivation and enzalutamide and performed immunofluorescence staining and western blotting to analyze their expression of AR and PSA.
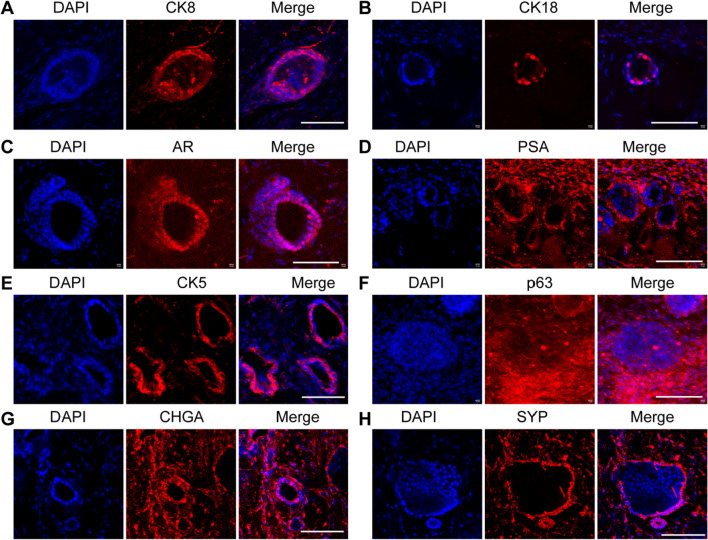
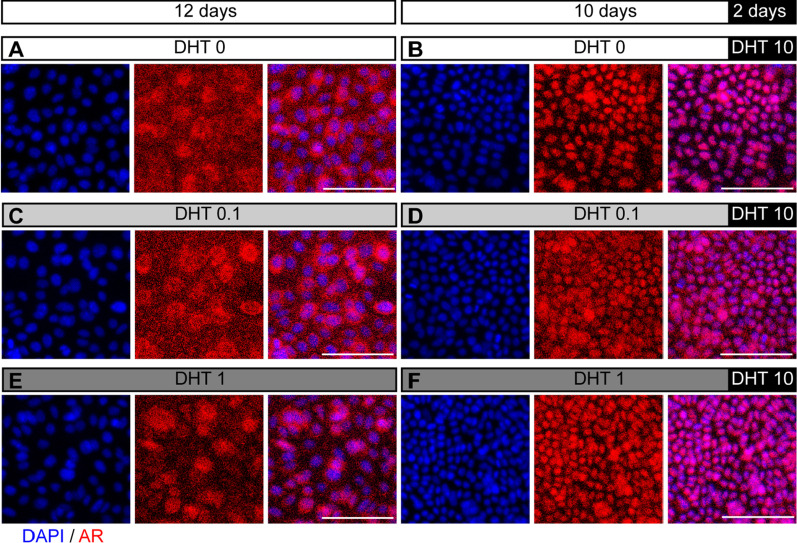
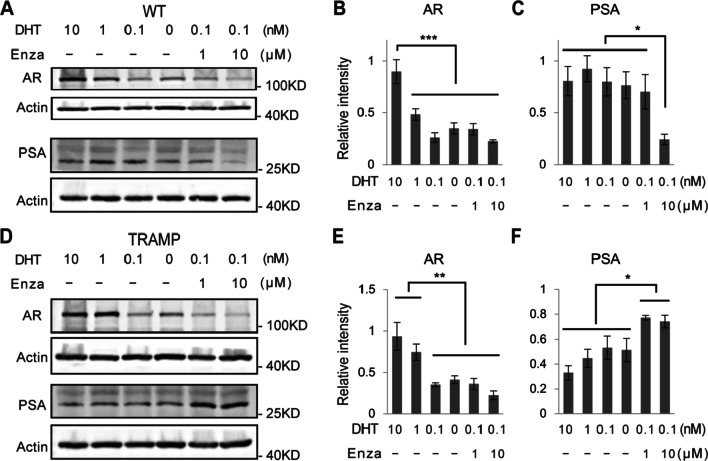
Results: We isolated a novel type of castration-resistant intermediate prostate stem cells (CRIPSCs) from adult WT and TRAMP mice. The mouse CRIPSCs proliferated rapidly in two-dimensional (2D) culture dishes and can be cultured for more than six months. The mouse CRIPSCs expressed luminal markers (AR, PSA, and Dsg4), basal markers (CK5 and p63), Psca, and the intermediate cell marker (Ivl). Transcriptomic analysis showed that the mouse CRIPSCs had upregulated signaling pathways related to cancer development and drug resistance. In the long-term culture, TRAMP CRIPSCs had higher expression of the genes related to stem cells and cancers than WT mice. Both WT and TRAMP CRIPSCs formed organoids in Matrigel. WT CRIPSCs did not form prostate tissues when transplanted in vivo without urogenital sinus mesenchyme (UGM) cells. In contrast, TRAMP CRIPSCs formed prostate ducts in NOG mice without UGM cells and differentiated into luminal, basal, and neuroendocrine cells. Androgens regulated AR translocation between the nucleus and cytoplasm in the mouse CRIPSCs. Treatment of androgen deprivation (ADT) and enzalutamide reduced AR expression in WT and TRAMP CRIPSCs; however, this treatment promoted PSA expression in TRAMP, while not WT CRIPSCs, similar to the clinical observations of CRPC.
Conclusions: Our study established a method for isolating and expanding mouse CRIPSCs in 2D culture dishes. Mouse CRIPSCs had markers of basal and luminal cells, including AR and PSA, and can differentiate into prostate organoids and tissues. TRAMP CRIPSCs had elevated PSA expression upon ADT and enzalutamide treatment. Our method can be translated into clinical settings for CRPC precision medicine.
Keywords: Androgen deprivation; Androgen receptor; Enzalutamide; Intermediate cell; Prostate-specific antigen; Stem cell; castration-resistant prostate cancer.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
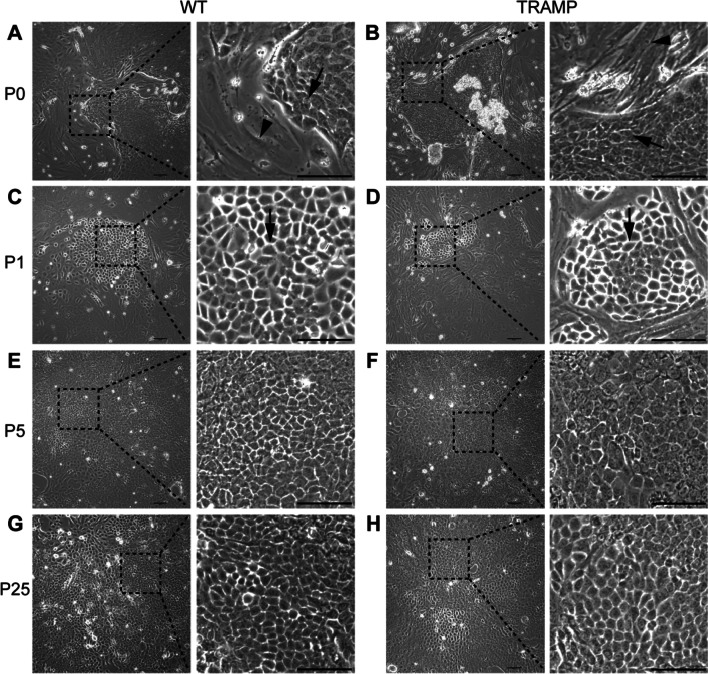
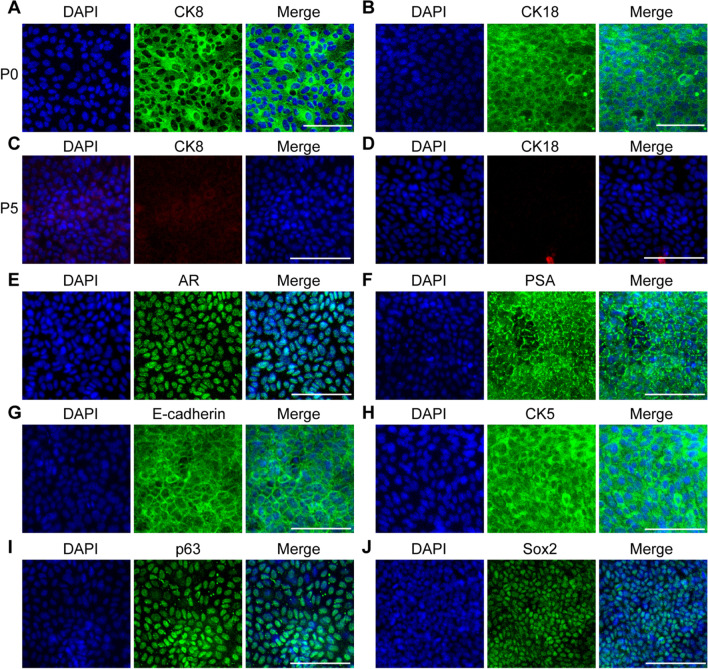
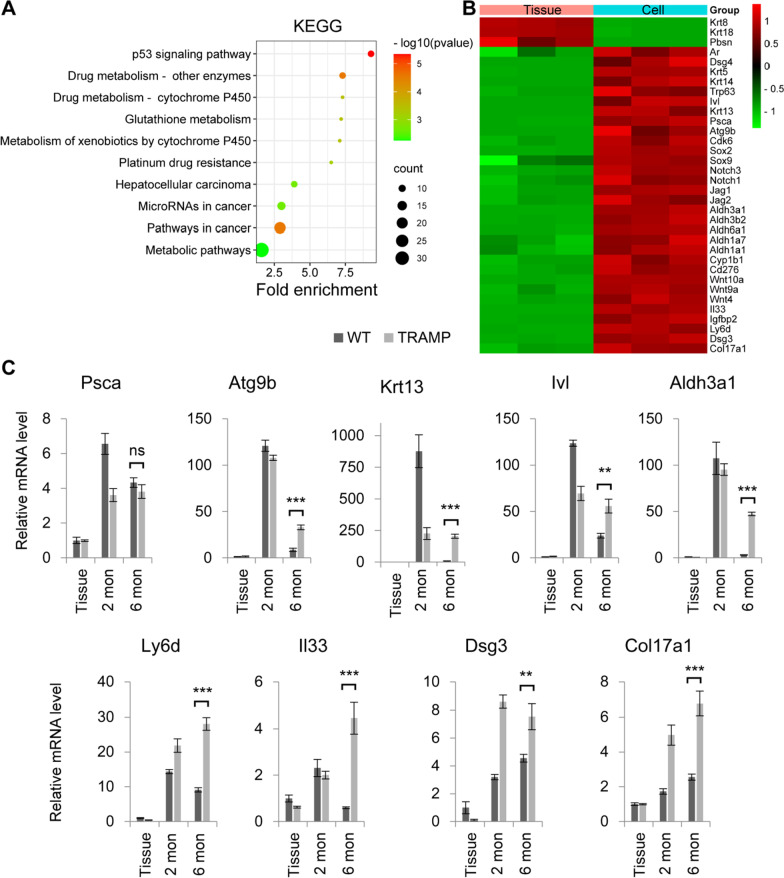
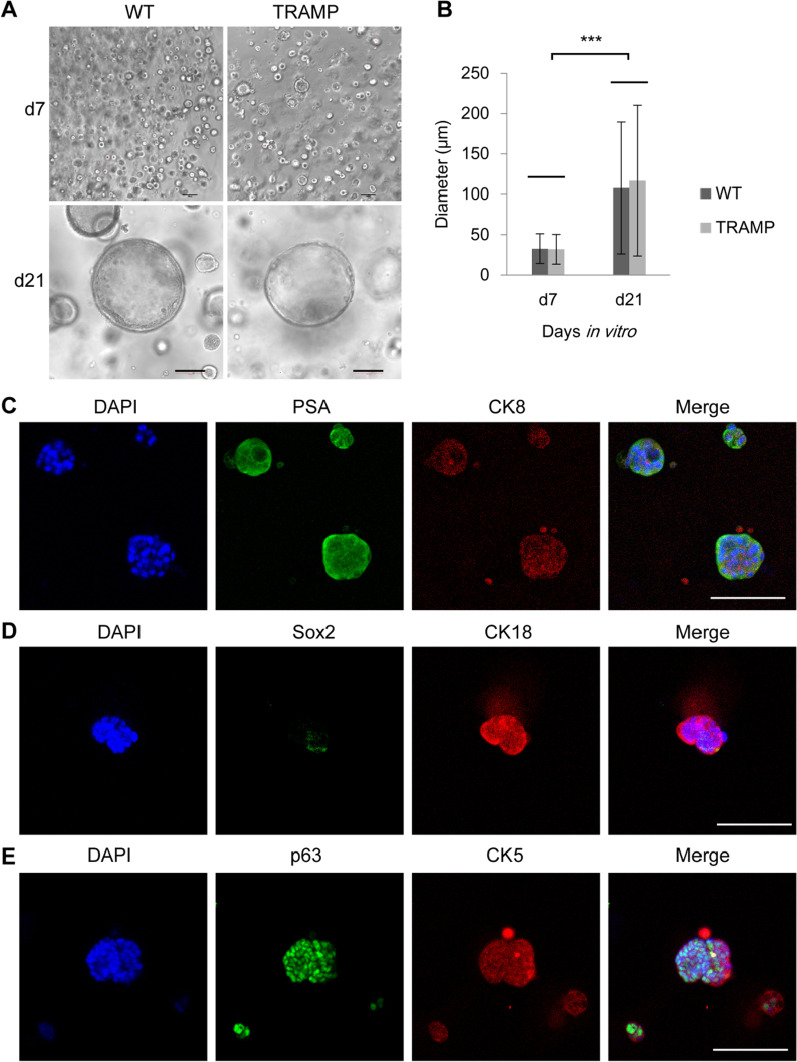
Similar articles
-
Human intermediate prostate cancer stem cells contribute to the initiation and development of prostate adenocarcinoma.Stem Cell Res Ther. 2024 Sep 11;15(1):296. doi: 10.1186/s13287-024-03917-8. Stem Cell Res Ther. 2024. PMID: 39256886 Free PMC article.
-
Androgen receptors in hormone-dependent and castration-resistant prostate cancer.Pharmacol Ther. 2013 Dec;140(3):223-38. doi: 10.1016/j.pharmthera.2013.07.003. Epub 2013 Jul 13. Pharmacol Ther. 2013. PMID: 23859952 Review.
-
Abiraterone switches castration-resistant prostate cancer dependency from adrenal androgens towards androgen receptor variants and glucocorticoid receptor signalling.Prostate. 2022 Apr;82(5):505-516. doi: 10.1002/pros.24297. Epub 2022 Jan 17. Prostate. 2022. PMID: 35037287 Free PMC article.
-
IU1 and enzalutamide combination yields synergistic effects on castration-resistant prostate cancer.Prostate. 2023 Nov;83(15):1446-1457. doi: 10.1002/pros.24607. Epub 2023 Aug 7. Prostate. 2023. PMID: 37545197
-
Targeting the androgen receptor signaling pathway in advanced prostate cancer.Am J Health Syst Pharm. 2022 Jul 22;79(15):1224-1235. doi: 10.1093/ajhp/zxac105. Am J Health Syst Pharm. 2022. PMID: 35390118 Review.
Cited by
-
CNKSR2 interactome analysis indicates its association with the centrosome/microtubule system.Neural Regen Res. 2025 Aug 1;20(8):2420-2432. doi: 10.4103/NRR.NRR-D-23-01725. Epub 2024 May 17. Neural Regen Res. 2025. PMID: 39359098 Free PMC article.
-
Two-dimensional vascularized liver organoid on extracellular matrix with defined stiffness for modeling fibrotic and normal tissues.J Tissue Eng. 2024 Aug 10;15:20417314241268344. doi: 10.1177/20417314241268344. eCollection 2024 Jan-Dec. J Tissue Eng. 2024. PMID: 39130682 Free PMC article.
-
Cancer-associated fibroblasts and prostate cancer stem cells: crosstalk mechanisms and implications for disease progression.Front Cell Dev Biol. 2024 Jul 18;12:1412337. doi: 10.3389/fcell.2024.1412337. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 39092186 Free PMC article. Review.
-
Electrochemiluminescence immunosensor based on Cu3(PO4)2 hybrid nanoflowers as a novel luminophore for the sensitive detection of prostate-specific antigen.Mikrochim Acta. 2023 Sep 12;190(10):389. doi: 10.1007/s00604-023-05966-x. Mikrochim Acta. 2023. PMID: 37700114
-
Human intermediate prostate cancer stem cells contribute to the initiation and development of prostate adenocarcinoma.Stem Cell Res Ther. 2024 Sep 11;15(1):296. doi: 10.1186/s13287-024-03917-8. Stem Cell Res Ther. 2024. PMID: 39256886 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
