

A reactive oxygen species Ca2+ signalling pathway identified from a chemical screen for modifiers of sugar-activated circadian gene expression
- PMID: 35842791
- PMCID: PMC9804775
- DOI: 10.1111/nph.18380
A reactive oxygen species Ca2+ signalling pathway identified from a chemical screen for modifiers of sugar-activated circadian gene expression
Abstract
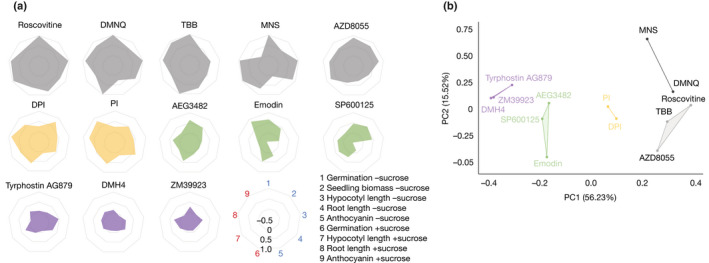
Sugars are essential metabolites for energy and anabolism that can also act as signals to regulate plant physiology and development. Experimental tools to disrupt major sugar signalling pathways are limited. We performed a chemical screen for modifiers of activation of circadian gene expression by sugars to discover pharmacological tools to investigate and manipulate plant sugar signalling. Using a library of commercially available bioactive compounds, we identified 75 confident hits that modified the response of a circadian luciferase reporter to sucrose in dark-adapted Arabidopsis thaliana seedlings. We validated the transcriptional effect on a subset of the hits and measured their effects on a range of sugar-dependent phenotypes for 13 of these chemicals. Chemicals were identified that appear to influence known and unknown sugar signalling pathways. Pentamidine isethionate was identified as a modifier of a sugar-activated Ca2+ signal that acts as a calmodulin inhibitor downstream of superoxide in a metabolic signalling pathway affecting circadian rhythms, primary metabolism and plant growth. Our data provide a resource of new experimental tools to manipulate plant sugar signalling and identify novel components of these pathways.
Keywords: Arabidopsis thaliana; LOPAC (Library of Pharmacologically Active Compounds); calcium; calmodulin; circadian clock; reactive oxygen species (ROS); sugar signalling; superoxide.
© 2022 The Authors. New Phytologist © 2022 New Phytologist Foundation.
Figures







References
-
- Astegno A, Bonza MC, Vallone R, la Verde V, D'Onofrio M, Luoni L, Molesini B, Dominici P. 2017. Arabidopsis calmodulin‐like protein CML36 is a calcium (Ca2+) sensor that interacts with the plasma membrane Ca2+‐ATPase isoform ACA8 and stimulates its activity. Journal of Biological Chemistry 292: 15049–15061. - PMC - PubMed
-
- Baena‐González E, Rolland F, Thevelein JM, Sheen J. 2007. A central integrator of transcription networks in plant stress and energy signalling. Nature 448: 938–942. - PubMed
-
- Belda‐Palazón B, Adamo M, Valerio C, Ferreira LJ, Confraria A, Reis‐Barata D, Rodrigues A, Meyer C, Rodriguez PL, Baena‐González E. 2020. A dual function of SnRK2 kinases in the regulation of SnRK1 and plant growth. Nature Plants 6: 1345–1353. - PubMed
-
- Bender KW, Rosenbaum DM, Vanderbeld B, Ubaid M, Snedden WA. 2013. The Arabidopsis calmodulin‐like protein, CML39, functions during early seedling establishment. The Plant Journal 76: 634–647. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
