

Neuronal Adenosine A1 Receptor is Critical for Olfactory Function but Unable to Attenuate Olfactory Dysfunction in Neuroinflammation
- PMID: 35846561
- PMCID: PMC9279574
- DOI: 10.3389/fncel.2022.912030
Neuronal Adenosine A1 Receptor is Critical for Olfactory Function but Unable to Attenuate Olfactory Dysfunction in Neuroinflammation
Abstract
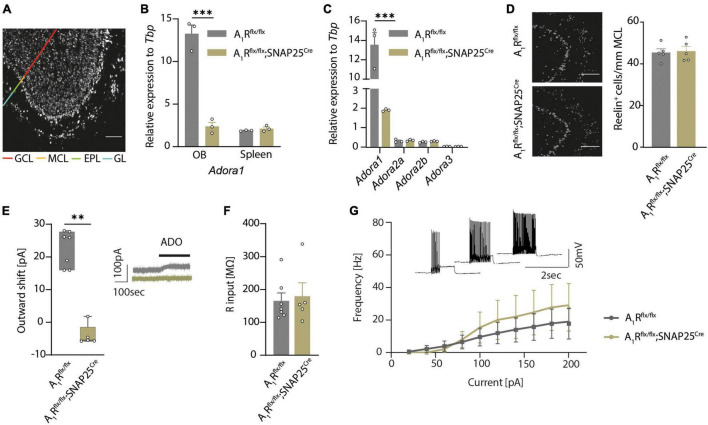
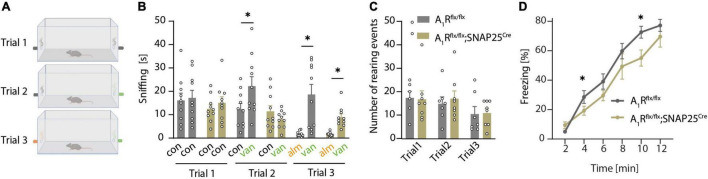
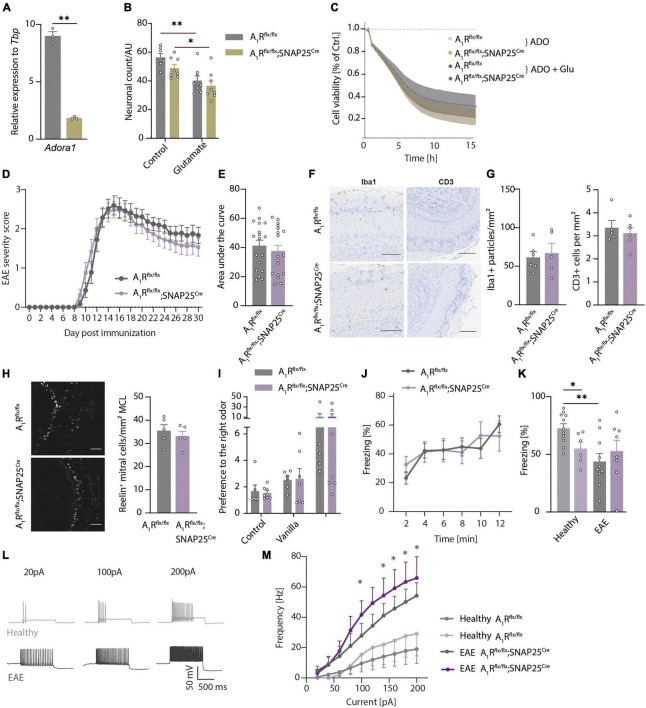
Adenine nucleotides, such as adenosine triphosphate (ATP), adenosine diphosphate (ADP), as well as the nucleoside adenosine are important modulators of neuronal function by engaging P1 and P2 purinergic receptors. In mitral cells, signaling of the G protein-coupled P1 receptor adenosine 1 receptor (A1R) affects the olfactory sensory pathway by regulating high voltage-activated calcium channels and two-pore domain potassium (K2P) channels. The inflammation of the central nervous system (CNS) impairs the olfactory function and gives rise to large amounts of extracellular ATP and adenosine, which act as pro-inflammatory and anti-inflammatory mediators, respectively. However, it is unclear whether neuronal A1R in the olfactory bulb modulates the sensory function and how this is impacted by inflammation. Here, we show that signaling via neuronal A1R is important for the physiological olfactory function, while it cannot counteract inflammation-induced hyperexcitability and olfactory deficit. Using neuron-specific A1R-deficient mice in patch-clamp recordings, we found that adenosine modulates spontaneous dendro-dendritic signaling in mitral and granule cells via A1R. Furthermore, neuronal A1R deficiency resulted in olfactory dysfunction in two separate olfactory tests. In mice with experimental autoimmune encephalomyelitis (EAE), we detected immune cell infiltration and microglia activation in the olfactory bulb as well as hyperexcitability of mitral cells and olfactory dysfunction. However, neuron-specific A1R activity was unable to attenuate glutamate excitotoxicity in the primary olfactory bulb neurons in vitro or EAE-induced olfactory dysfunction and disease severity in vivo. Together, we demonstrate that A1R modulates the dendro-dendritic inhibition (DDI) at the site of mitral and granule cells and impacts the processing of the olfactory sensory information, while A1R activity was unable to counteract inflammation-induced hyperexcitability.
Keywords: A1R; EAE; adenosine; mitral cells; neuroprotection; olfactory bulb; olfactory dysfunction; purinergic signaling.
Copyright © 2022 Schubert, Schulz, Träger, Plath, Omriouate, Rosenkranz, Morellini, Friese and Hirnet.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
Impaired olfactory performance and anxiety-like behavior in a rat model of multiple sclerosis are associated with enhanced adenosine signaling in the olfactory bulb via A1R, A2BR, and A3R.Front Cell Neurosci. 2024 Jul 30;18:1407975. doi: 10.3389/fncel.2024.1407975. eCollection 2024. Front Cell Neurosci. 2024. PMID: 39139401 Free PMC article.
-
Adenosine A1 receptor activates background potassium channels and modulates information processing in olfactory bulb mitral cells.J Physiol. 2018 Feb 15;596(4):717-733. doi: 10.1113/JP275503. Epub 2018 Jan 24. J Physiol. 2018. PMID: 29274133 Free PMC article.
-
Adenosine A1 Receptor-Mediated Attenuation of Reciprocal Dendro-Dendritic Inhibition in the Mouse Olfactory Bulb.Front Cell Neurosci. 2018 Jan 15;11:435. doi: 10.3389/fncel.2017.00435. eCollection 2017. Front Cell Neurosci. 2018. PMID: 29379418 Free PMC article.
-
The role of adenosine A1 receptor on immune cells.Inflamm Res. 2022 Nov;71(10-11):1203-1212. doi: 10.1007/s00011-022-01607-w. Epub 2022 Sep 5. Inflamm Res. 2022. PMID: 36064866 Review.
-
Purinergic neuron-glia interactions in sensory systems.Pflugers Arch. 2014 Oct;466(10):1859-72. doi: 10.1007/s00424-014-1510-6. Epub 2014 Apr 6. Pflugers Arch. 2014. PMID: 24705940 Review.
Cited by
-
Ion Channels in Odor Information Processing of Neural Circuits of the Vertebrate Olfactory Bulb.Int J Mol Sci. 2024 Dec 10;25(24):13259. doi: 10.3390/ijms252413259. Int J Mol Sci. 2024. PMID: 39769024 Free PMC article. Review.
-
Increased Purinergic Signaling in Human Dental Pulps With Inflammatory Pain is Sex-Dependent.J Pain. 2024 Apr;25(4):1039-1058. doi: 10.1016/j.jpain.2023.10.026. Epub 2023 Nov 11. J Pain. 2024. PMID: 37956743 Free PMC article.
-
Impaired olfactory performance and anxiety-like behavior in a rat model of multiple sclerosis are associated with enhanced adenosine signaling in the olfactory bulb via A1R, A2BR, and A3R.Front Cell Neurosci. 2024 Jul 30;18:1407975. doi: 10.3389/fncel.2024.1407975. eCollection 2024. Front Cell Neurosci. 2024. PMID: 39139401 Free PMC article.
-
A2A adenosine receptor-driven cAMP signaling in olfactory bulb astrocytes is unaffected in experimental autoimmune encephalomyelitis.Front Immunol. 2023 Nov 23;14:1273837. doi: 10.3389/fimmu.2023.1273837. eCollection 2023. Front Immunol. 2023. PMID: 38077336 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases