

Respiratory syncytial virus disrupts the airway epithelial barrier by decreasing cortactin and destabilizing F-actin
- PMID: 35848790
- PMCID: PMC9481929
- DOI: 10.1242/jcs.259871
Respiratory syncytial virus disrupts the airway epithelial barrier by decreasing cortactin and destabilizing F-actin
Abstract
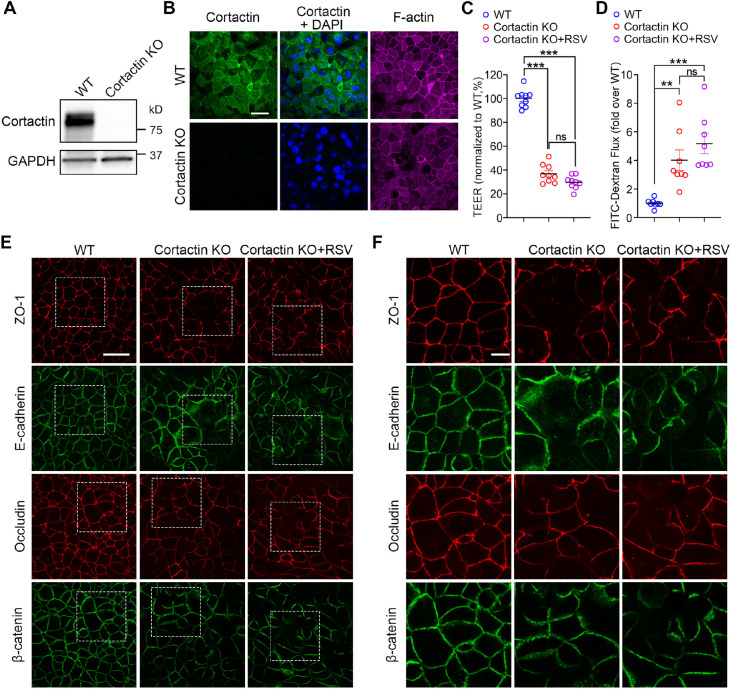
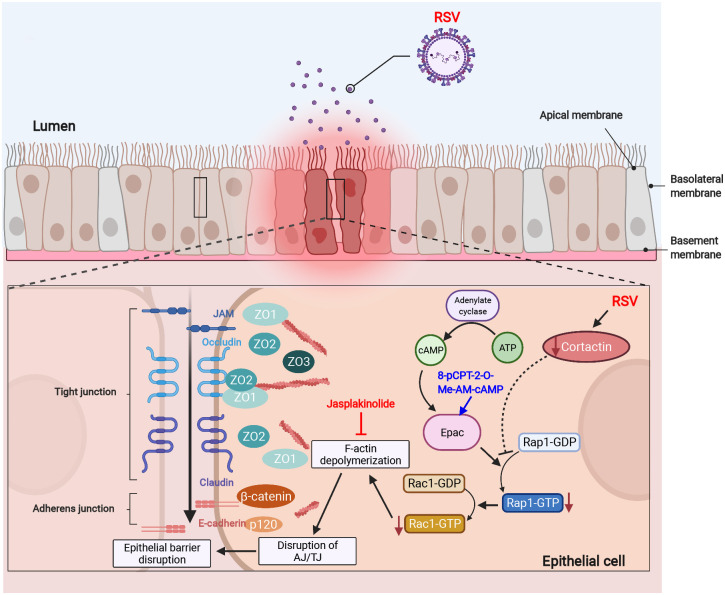
Respiratory syncytial virus (RSV) infection is the leading cause of acute lower respiratory tract infection in young children worldwide. Our group recently revealed that RSV infection disrupts the airway epithelial barrier in vitro and in vivo. However, the underlying molecular pathways were still elusive. Here, we report the critical roles of the filamentous actin (F-actin) network and actin-binding protein cortactin in RSV infection. We found that RSV infection causes F-actin depolymerization in 16HBE cells, and that stabilizing the F-actin network in infected cells reverses the epithelial barrier disruption. RSV infection also leads to significantly decreased cortactin in vitro and in vivo. Cortactin-knockout 16HBE cells presented barrier dysfunction, whereas overexpression of cortactin protected the epithelial barrier against RSV. The activity of Rap1 (which has Rap1A and Rap1B forms), one downstream target of cortactin, declined after RSV infection as well as in cortactin-knockout cells. Moreover, activating Rap1 attenuated RSV-induced epithelial barrier disruption. Our study proposes a key mechanism in which RSV disrupts the airway epithelial barrier via attenuating cortactin expression and destabilizing the F-actin network. The identified pathways will provide new targets for therapeutic intervention toward RSV-related disease. This article has an associated First Person interview with the first author of the paper.
Keywords: Airway epithelial cells; Cortactin; Epithelial barrier; F-actin; RSV; Rap1 GTPase; Respiratory syncytial virus.
© 2022. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures






Similar articles
-
Vitamin D3 protects against respiratory syncytial virus-induced barrier dysfunction in airway epithelial cells via PKA signaling pathway.Eur J Cell Biol. 2023 Sep;102(3):151336. doi: 10.1016/j.ejcb.2023.151336. Epub 2023 Jun 22. Eur J Cell Biol. 2023. PMID: 37354621 Free PMC article.
-
Disruption of the airway epithelial barrier in a murine model of respiratory syncytial virus infection.Am J Physiol Lung Cell Mol Physiol. 2019 Feb 1;316(2):L358-L368. doi: 10.1152/ajplung.00345.2018. Epub 2018 Nov 29. Am J Physiol Lung Cell Mol Physiol. 2019. PMID: 30489157 Free PMC article.
-
cAMP-dependent activation of protein kinase A attenuates respiratory syncytial virus-induced human airway epithelial barrier disruption.PLoS One. 2017 Jul 31;12(7):e0181876. doi: 10.1371/journal.pone.0181876. eCollection 2017. PLoS One. 2017. PMID: 28759570 Free PMC article.
-
New insights on the viral and host factors contributing to the airway pathogenesis caused by the respiratory syncytial virus.Crit Rev Microbiol. 2016 Sep;42(5):800-12. doi: 10.3109/1040841X.2015.1055711. Epub 2015 Jun 29. Crit Rev Microbiol. 2016. PMID: 26119025 Review.
-
Contribution of neuroimmune mechanisms to airway inflammation and remodeling during and after respiratory syncytial virus infection.Pediatr Infect Dis J. 2003 Feb;22(2 Suppl):S66-74; discussion S74-5. doi: 10.1097/01.inf.0000053888.67311.1d. Pediatr Infect Dis J. 2003. PMID: 12671455 Review.
Cited by
-
Electronic cigarette exposure disrupts airway epithelial barrier function and exacerbates viral infection.Am J Physiol Lung Cell Mol Physiol. 2023 Nov 1;325(5):L580-L593. doi: 10.1152/ajplung.00135.2023. Epub 2023 Sep 12. Am J Physiol Lung Cell Mol Physiol. 2023. PMID: 37698113 Free PMC article.
-
Infection of Primary Nasal Epithelial Cells Grown at an Air-Liquid Interface to Characterize Human Coronavirus-Host Interactions.J Vis Exp. 2023 Sep 22;(199):10.3791/64868. doi: 10.3791/64868. J Vis Exp. 2023. PMID: 37811957 Free PMC article.
-
Heavy metal-induced disruption of the autophagy-lysosomal pathway: implications for aging and neurodegenerative disorders.Biometals. 2025 Apr;38(2):371-417. doi: 10.1007/s10534-025-00665-x. Epub 2025 Feb 17. Biometals. 2025. PMID: 39960543 Review.
-
The impact of electronic cigarettes on airway epithelial barrier integrity in preclinical mouse model.Am J Physiol Lung Cell Mol Physiol. 2025 Apr 1;328(4):L564-L570. doi: 10.1152/ajplung.00408.2024. Epub 2025 Mar 10. Am J Physiol Lung Cell Mol Physiol. 2025. PMID: 40059636 Free PMC article.
-
Inhibiting F-Actin Polymerization Impairs the Internalization of Moraxella catarrhalis.Microorganisms. 2024 Jan 30;12(2):291. doi: 10.3390/microorganisms12020291. Microorganisms. 2024. PMID: 38399695 Free PMC article.
References
-
- Adamson, R. H., Ly, J. C., Sarai, R. K., Lenz, J. F., Altangerel, A., Drenckhahn, D. and Curry, F. E. (2008). Epac/Rap1 pathway regulates microvascular hyperpermeability induced by PAF in rat mesentery. Am. J. Physiol. Heart Circ. Physiol. 294, H1188-H1196. 10.1152/ajpheart.00937.2007 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
