

Tissue Homeostasis and Non-Homeostasis: From Cell Life Cycles to Organ States
- PMID: 35850152
- PMCID: PMC10182861
- DOI: 10.1146/annurev-cellbio-120420-114855
Tissue Homeostasis and Non-Homeostasis: From Cell Life Cycles to Organ States
Abstract
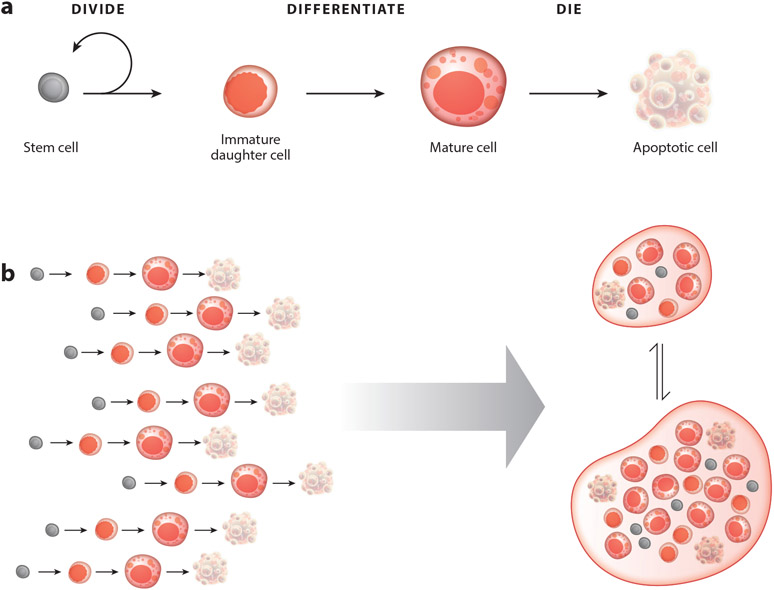
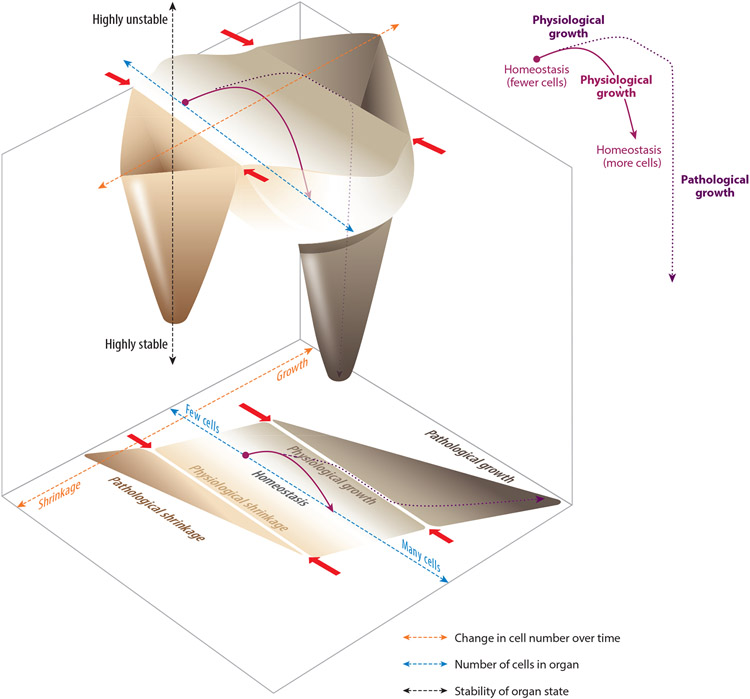
Although tissue homeostasis-the steady state-implies stability, our organs are in a state of continual, large-scale cellular flux. This flux underpins an organ's ability to homeostatically renew, to non-homeostatically resize upon altered functional demand, and to return to homeostasis after resizing or injury-in other words, to be dynamic. Here, I examine the basic unit of organ-scale cell dynamics: the cellular life cycle of birth, differentiation, and death. Focusing on epithelial organs, I discuss how spatial patterns and temporal kinetics of life cycle stages depend upon lineage organization and tissue architecture. I review how signaling between stages coordinates life cycle dynamics to enforce homeostasis, and I highlight how particular stages are transiently unbalanced to drive organ resizing or repair. Finally, I offer that considering organs as a collective of not cells but rather cell life cycles provides a powerful vantage for deciphering homeostatic and non-homeostatic tissue states.
Keywords: adaptation; differentiation; extrusion; feedback; stem cells; tissue homeostasis.
Figures
References
-
- Altmann GG. 1972. Influence of starvation and refeeding on mucosal size and epithelial renewal in the rat small intestine. Am. J. Anat 133(4):391–400 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
