

Engineering circular RNA for enhanced protein production
- PMID: 35851375
- PMCID: PMC9931579
- DOI: 10.1038/s41587-022-01393-0
Engineering circular RNA for enhanced protein production
Erratum in
-
Author Correction: Engineering circular RNA for enhanced protein production.Nat Biotechnol. 2023 Feb;41(2):293. doi: 10.1038/s41587-022-01472-2. Nat Biotechnol. 2023. PMID: 35978134 Free PMC article. No abstract available.
Abstract
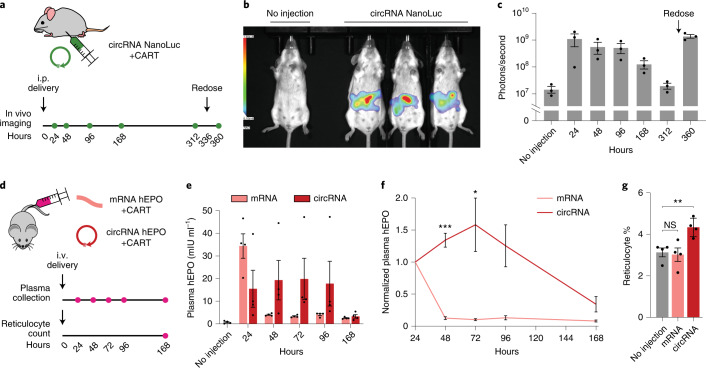
Circular RNAs (circRNAs) are stable and prevalent RNAs in eukaryotic cells that arise from back-splicing. Synthetic circRNAs and some endogenous circRNAs can encode proteins, raising the promise of circRNA as a platform for gene expression. In this study, we developed a systematic approach for rapid assembly and testing of features that affect protein production from synthetic circRNAs. To maximize circRNA translation, we optimized five elements: vector topology, 5' and 3' untranslated regions, internal ribosome entry sites and synthetic aptamers recruiting translation initiation machinery. Together, these design principles improve circRNA protein yields by several hundred-fold, provide increased translation over messenger RNA in vitro, provide more durable translation in vivo and are generalizable across multiple transgenes.
© 2022. The Author(s).
Conflict of interest statement
Stanford University has filed patent applications based on this work in which R.C., S.K.W., L.A. and H.Y.C. are named as inventors. R.C. is an advisor to Circ Bio. H.Y.C. is a co-founder of Accent Therapeutics, Boundless Bio, Cartography Biosciences and Circ Bio and an advisor to 10x Genomics, Arsenal Biosciences and Spring Discovery. P.A.W. is a co-founder of BryoLogyx and N1 Life and an advisor to BryoLogyx, N1 Life, Synaptogenix, Cytokinetics, Evonik, Super Trans Medical, Ativo and Vault Pharma. The remaining authors declare no competing interests.
Figures
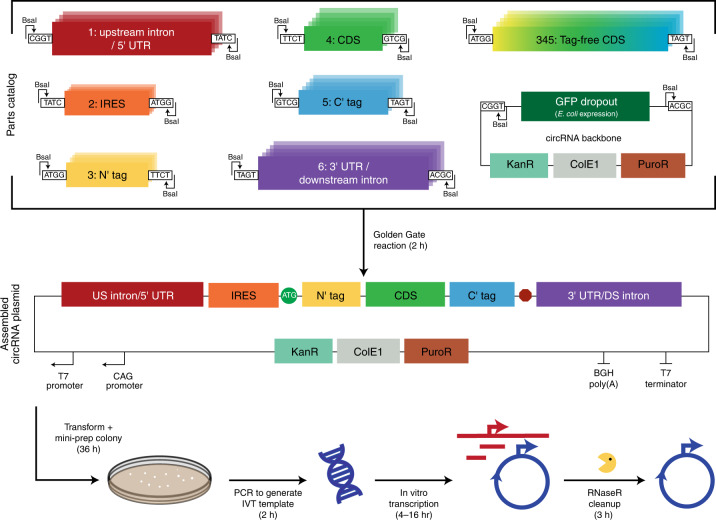
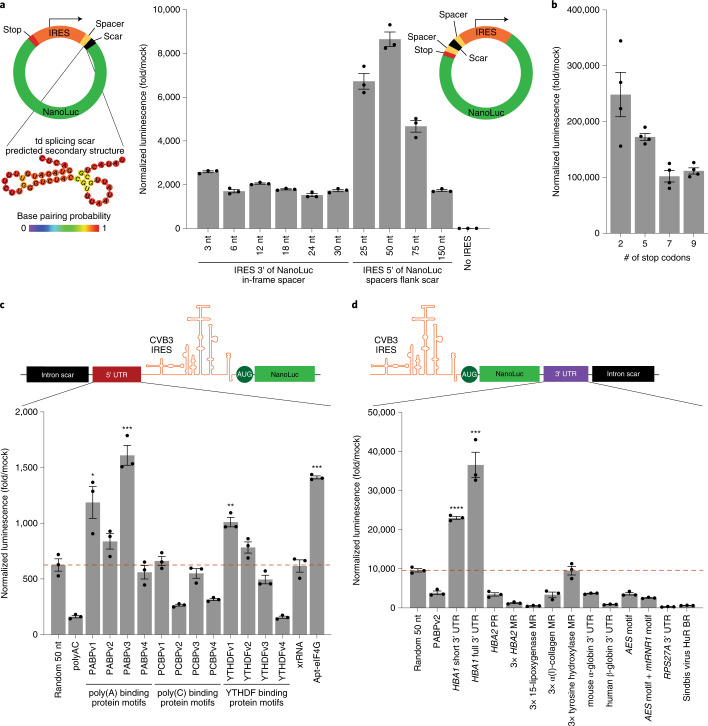

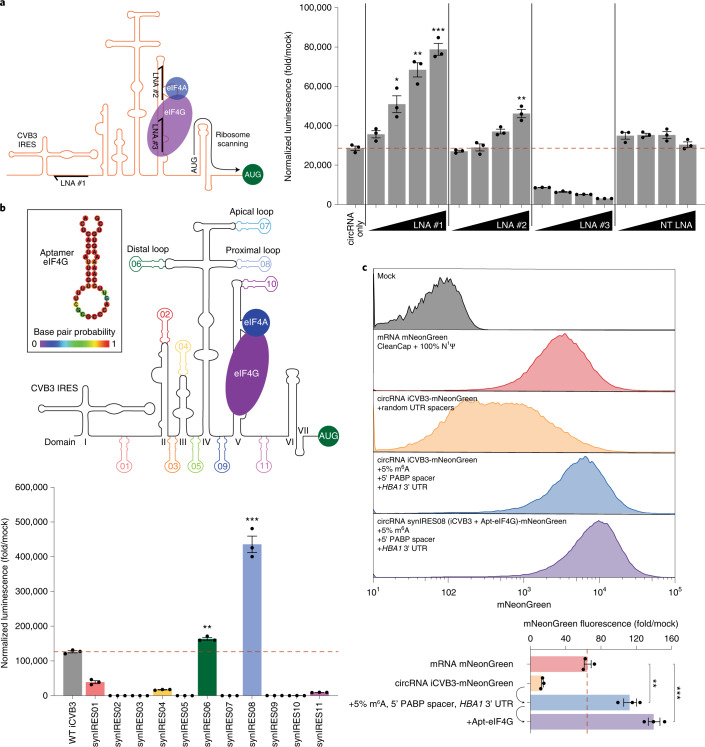
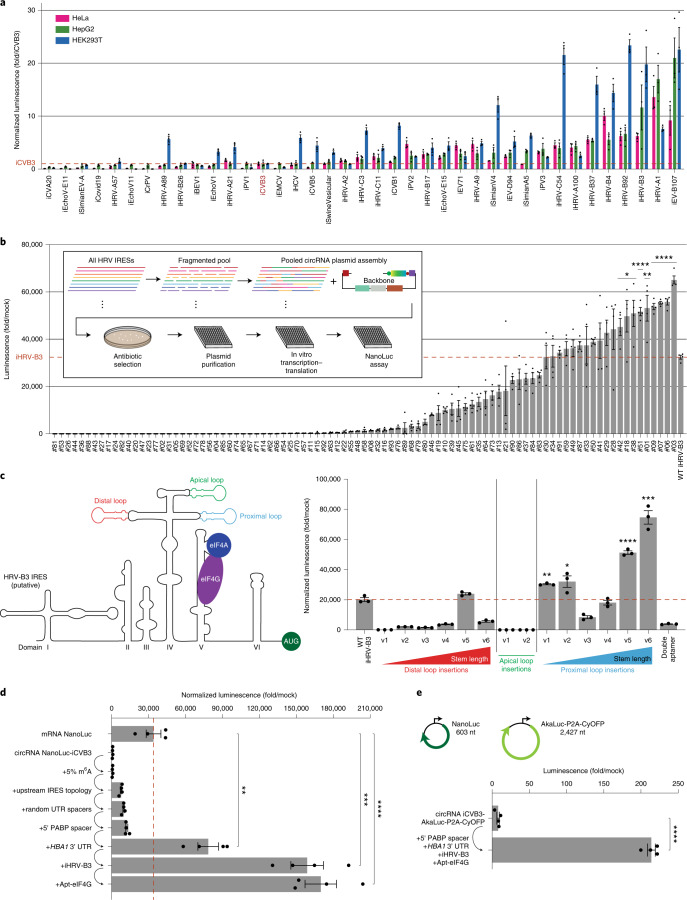
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
