

A new mechanism of fibronectin fibril assembly revealed by live imaging and super-resolution microscopy
- PMID: 35851804
- PMCID: PMC9481930
- DOI: 10.1242/jcs.260120
A new mechanism of fibronectin fibril assembly revealed by live imaging and super-resolution microscopy
Abstract
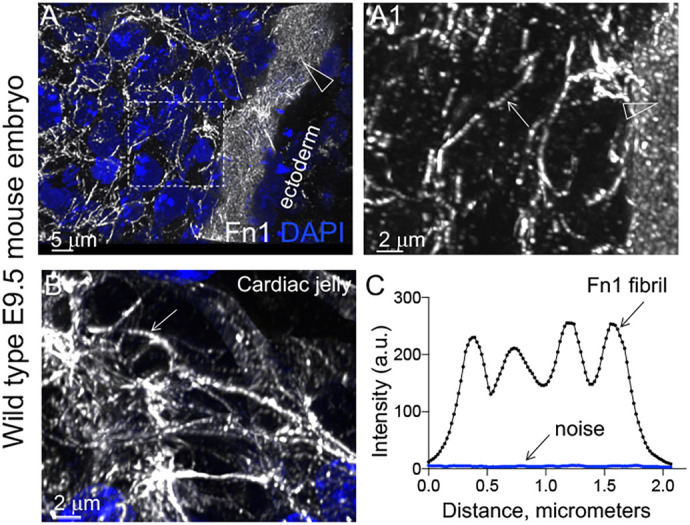
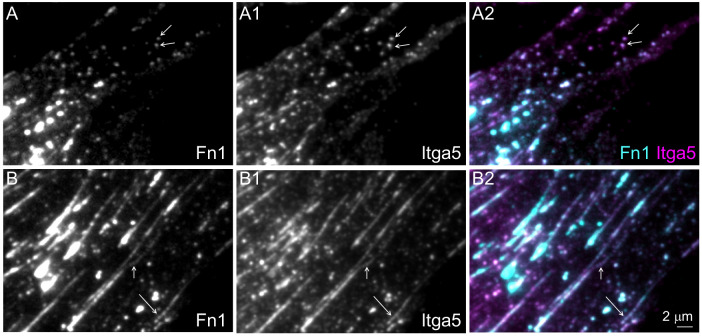
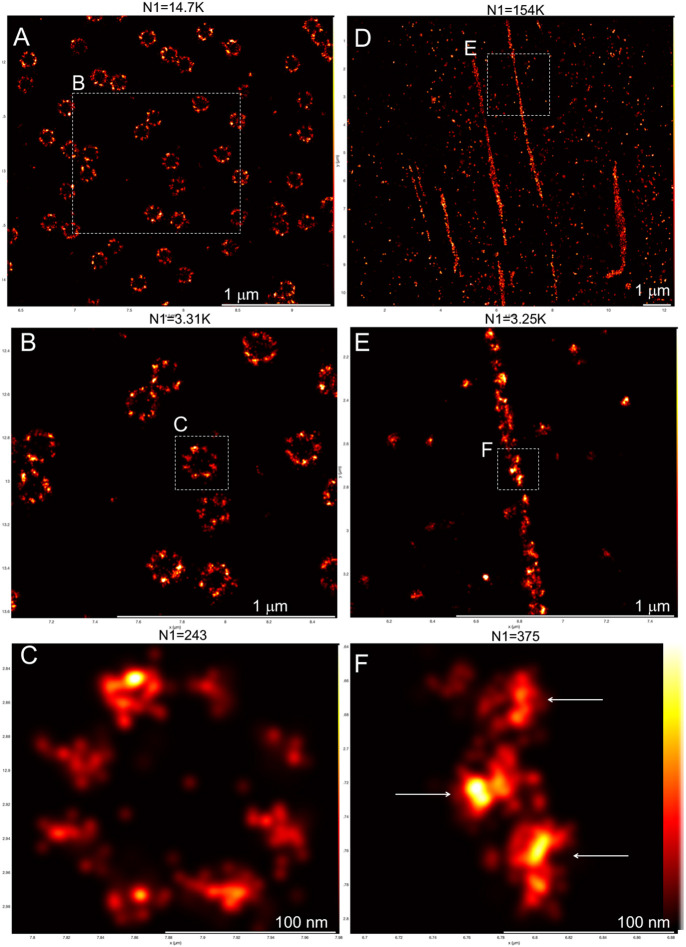
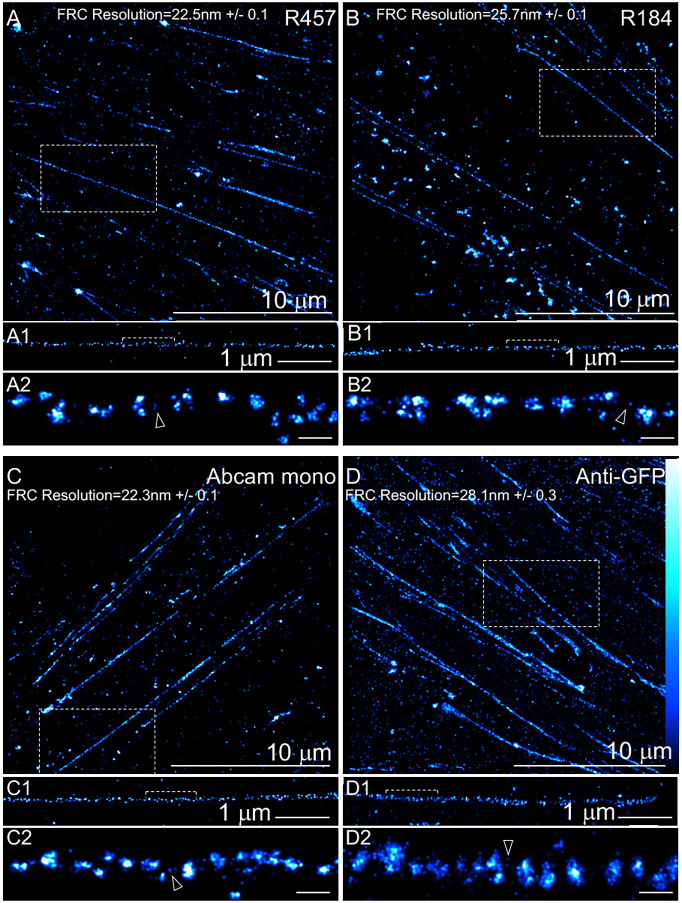
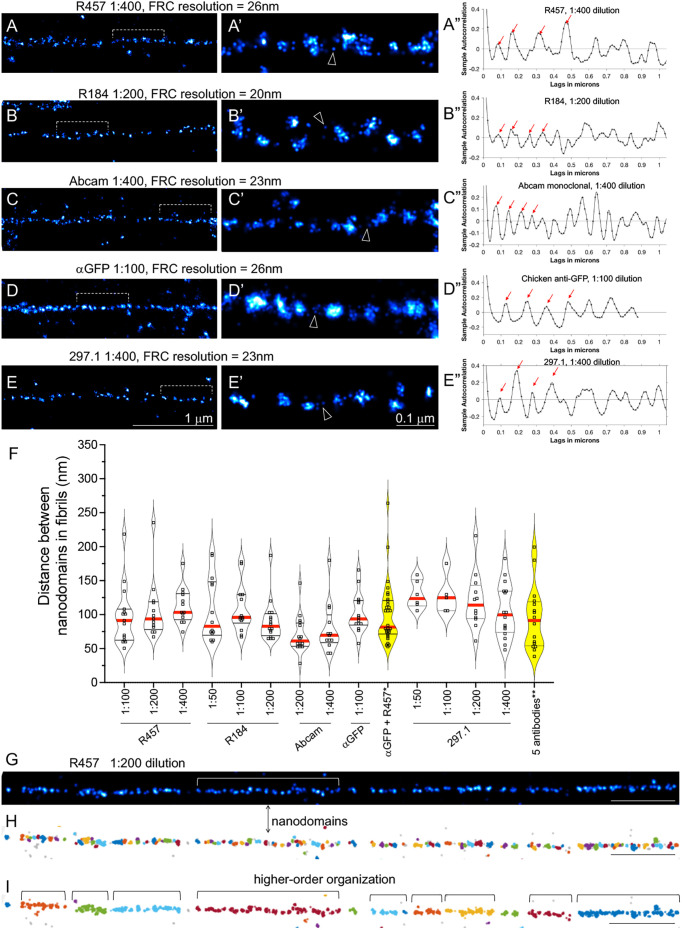
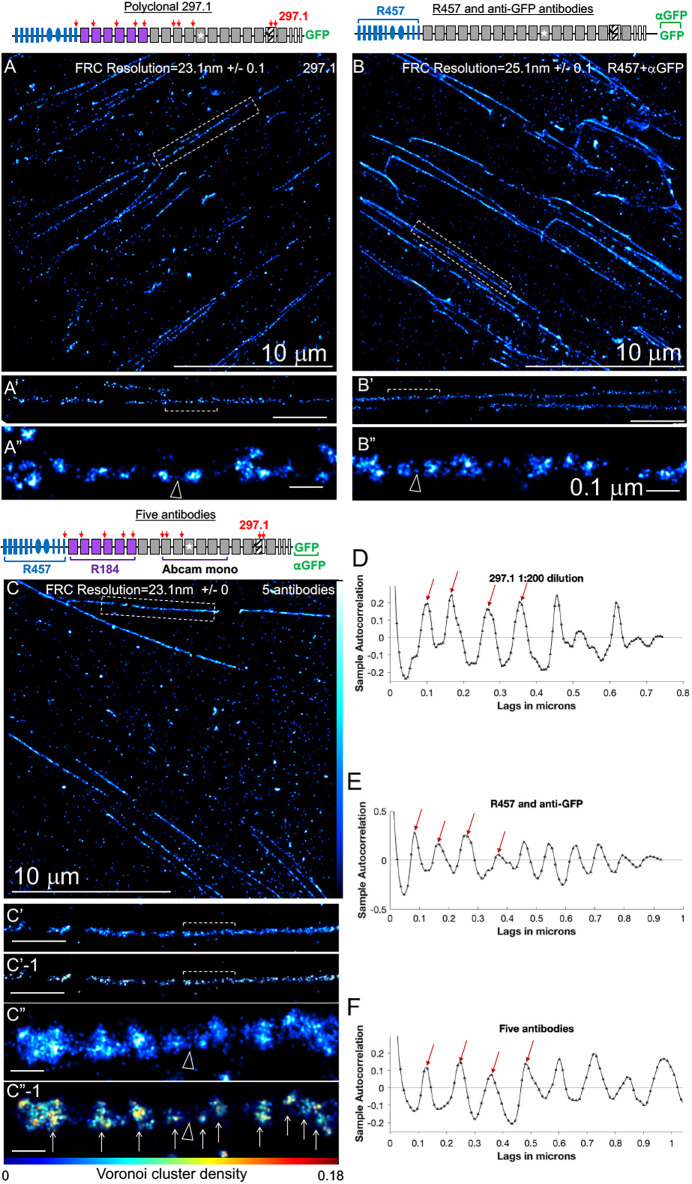
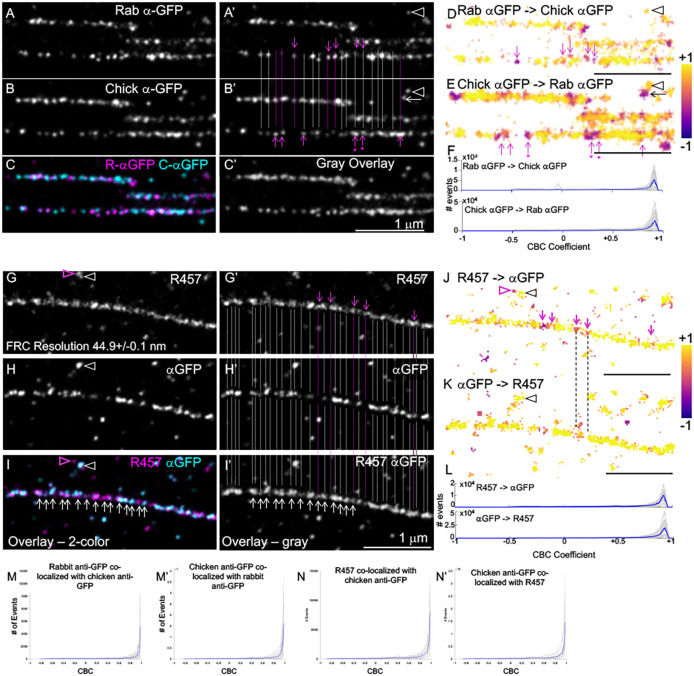
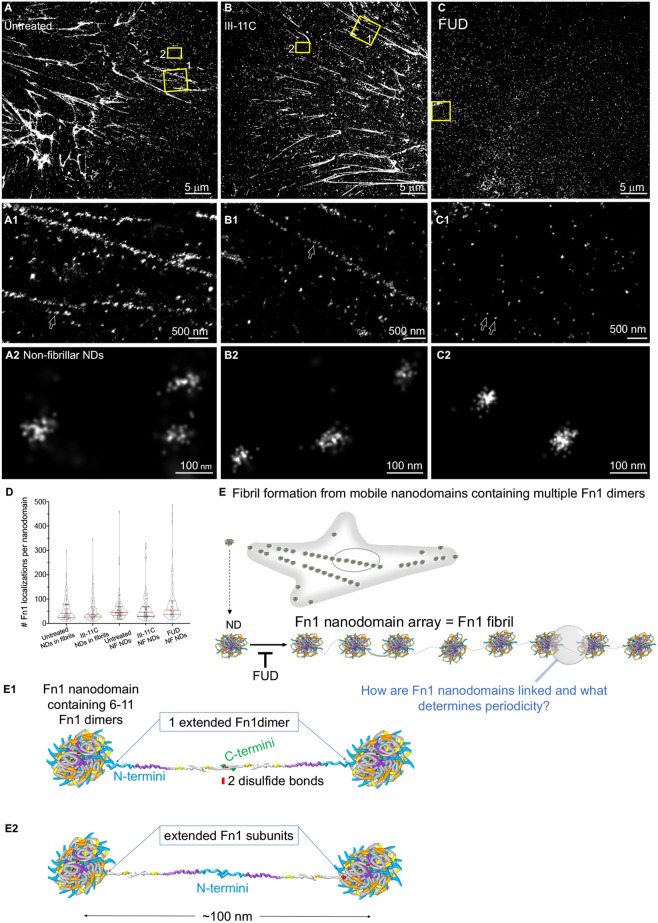
Fibronectin (Fn1) fibrils have long been viewed as continuous fibers composed of extended, periodically aligned Fn1 molecules. However, our live-imaging and single-molecule localization microscopy data are inconsistent with this traditional view and show that Fn1 fibrils are composed of roughly spherical nanodomains containing six to eleven Fn1 dimers. As they move toward the cell center, Fn1 nanodomains become organized into linear arrays, in which nanodomains are spaced with an average periodicity of 105±17 nm. Periodical Fn1 nanodomain arrays can be visualized between cells in culture and within tissues; they are resistant to deoxycholate treatment and retain nanodomain periodicity in the absence of cells. The nanodomain periodicity in fibrils remained constant when probed with antibodies recognizing distinct Fn1 epitopes or combinations of antibodies recognizing epitopes spanning the length of Fn1. Treatment with FUD, a peptide that binds the Fn1 N-terminus and disrupts Fn1 fibrillogenesis, blocked the organization of Fn1 nanodomains into periodical arrays. These studies establish a new paradigm of Fn1 fibrillogenesis. This article has an associated First Person interview with the first author of the paper.
Keywords: Fibril; Fibrillogenesis; Fibronectin; Live imaging; Super-resolution microscopy.
© 2022. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
Similar articles
-
Disruption of fibronectin fibrillogenesis affects intraocular pressure (IOP) in BALB/cJ mice.PLoS One. 2020 Aug 21;15(8):e0237932. doi: 10.1371/journal.pone.0237932. eCollection 2020. PLoS One. 2020. PMID: 32822410 Free PMC article.
-
PEGylated pUR4/FUD peptide inhibitor of fibronectin fibrillogenesis decreases fibrosis in murine Unilateral Ureteral Obstruction model of kidney disease.PLoS One. 2018 Oct 24;13(10):e0205360. doi: 10.1371/journal.pone.0205360. eCollection 2018. PLoS One. 2018. PMID: 30356276 Free PMC article.
-
Fibronectin Extra Domains tune cellular responses and confer topographically distinct features to fibril networks.J Cell Sci. 2021 Feb 24;134(4):jcs252957. doi: 10.1242/jcs.252957. J Cell Sci. 2021. PMID: 33526715
-
Collagen fibrillogenesis: fibronectin, integrins, and minor collagens as organizers and nucleators.Curr Opin Cell Biol. 2008 Oct;20(5):495-501. doi: 10.1016/j.ceb.2008.06.008. Epub 2008 Jul 30. Curr Opin Cell Biol. 2008. PMID: 18640274 Free PMC article. Review.
-
Fibronectin fibrillogenesis: a paradigm for extracellular matrix assembly.Curr Opin Cell Biol. 1999 Oct;11(5):622-7. doi: 10.1016/s0955-0674(99)00017-4. Curr Opin Cell Biol. 1999. PMID: 10508649 Review.
Cited by
-
Initiation of fibronectin fibrillogenesis is an enzyme-dependent process.Cell Rep. 2023 May 30;42(5):112473. doi: 10.1016/j.celrep.2023.112473. Epub 2023 May 5. Cell Rep. 2023. PMID: 37148241 Free PMC article.
-
Fibrillar adhesion dynamics govern the timescales of nuclear mechano-response via the vimentin cytoskeleton.bioRxiv [Preprint]. 2023 Nov 11:2023.11.08.566191. doi: 10.1101/2023.11.08.566191. bioRxiv. 2023. PMID: 37986921 Free PMC article. Preprint.
-
EDA Fibronectin Microarchitecture and YAP Translocation During Wound Closure.bioRxiv [Preprint]. 2024 Sep 25:2024.09.23.614581. doi: 10.1101/2024.09.23.614581. bioRxiv. 2024. Update in: ACS Biomater Sci Eng. 2025 Apr 14;11(4):2249-2262. doi: 10.1021/acsbiomaterials.4c02019. PMID: 39386582 Free PMC article. Updated. Preprint.
-
Live imaging of Fibronectin 1a-mNeonGreen and Fibronectin 1b-mCherry knock-in alleles during early zebrafish development.Cells Dev. 2024 Mar;177:203900. doi: 10.1016/j.cdev.2024.203900. Epub 2024 Jan 12. Cells Dev. 2024. PMID: 38218338 Free PMC article.
-
Mesodermal fibronectin controls cell shape, polarity, and mechanotransduction in the second heart field during cardiac outflow tract development.Dev Cell. 2025 Jan 6;60(1):62-84.e7. doi: 10.1016/j.devcel.2024.09.017. Epub 2024 Oct 15. Dev Cell. 2025. PMID: 39413783
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
