

Microglia deficiency accelerates prion disease but does not enhance prion accumulation in the brain
- PMID: 35852018
- PMCID: PMC9544114
- DOI: 10.1002/glia.24244
Microglia deficiency accelerates prion disease but does not enhance prion accumulation in the brain
Abstract
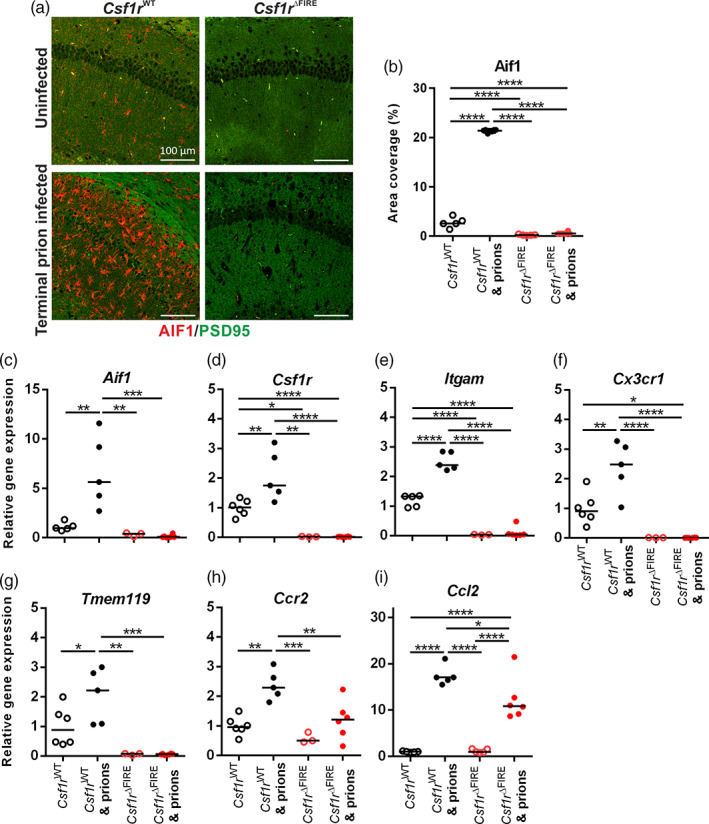
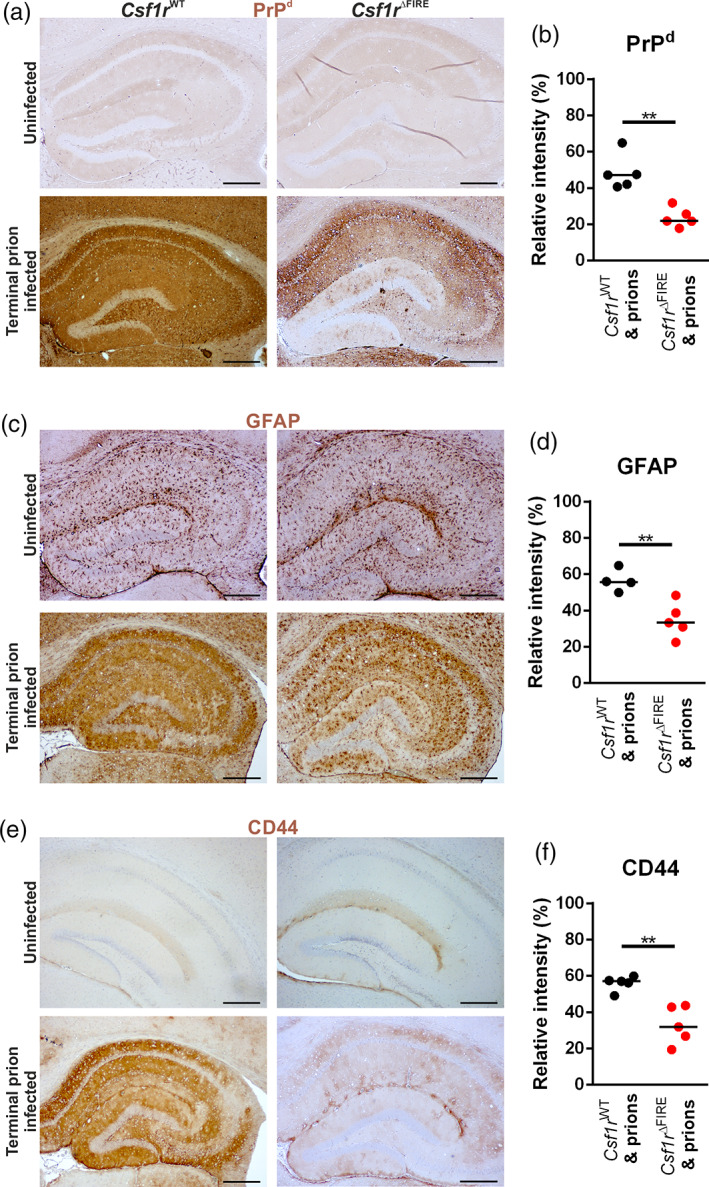
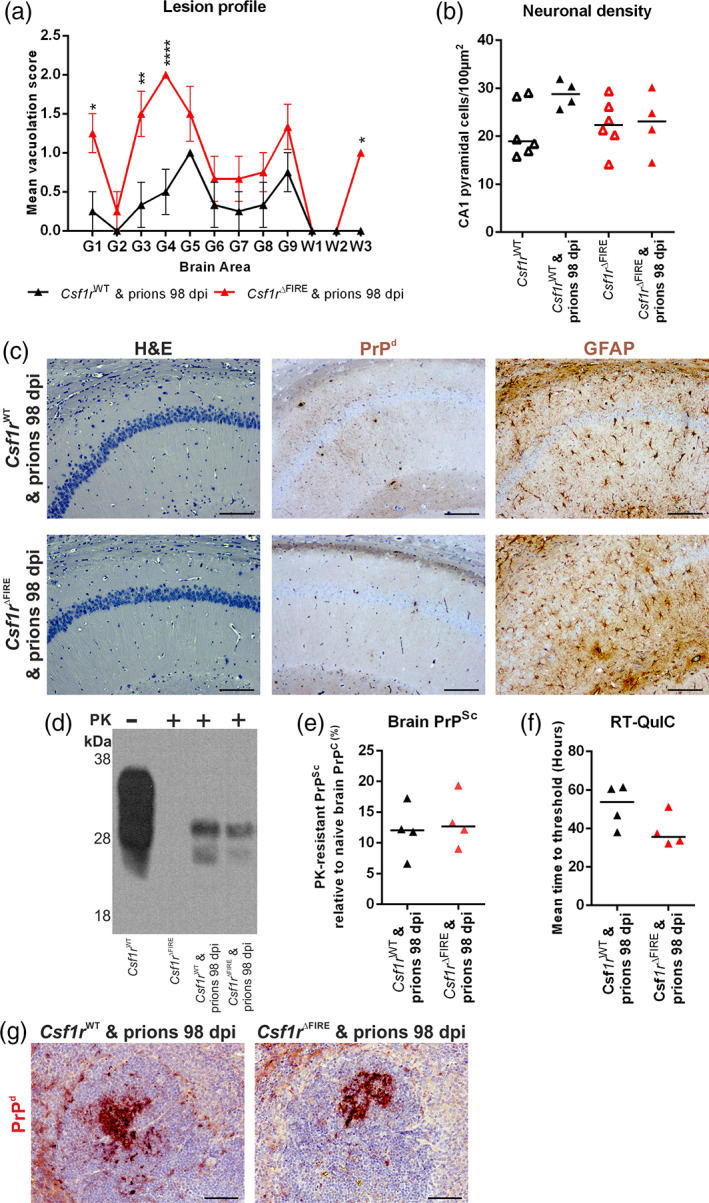
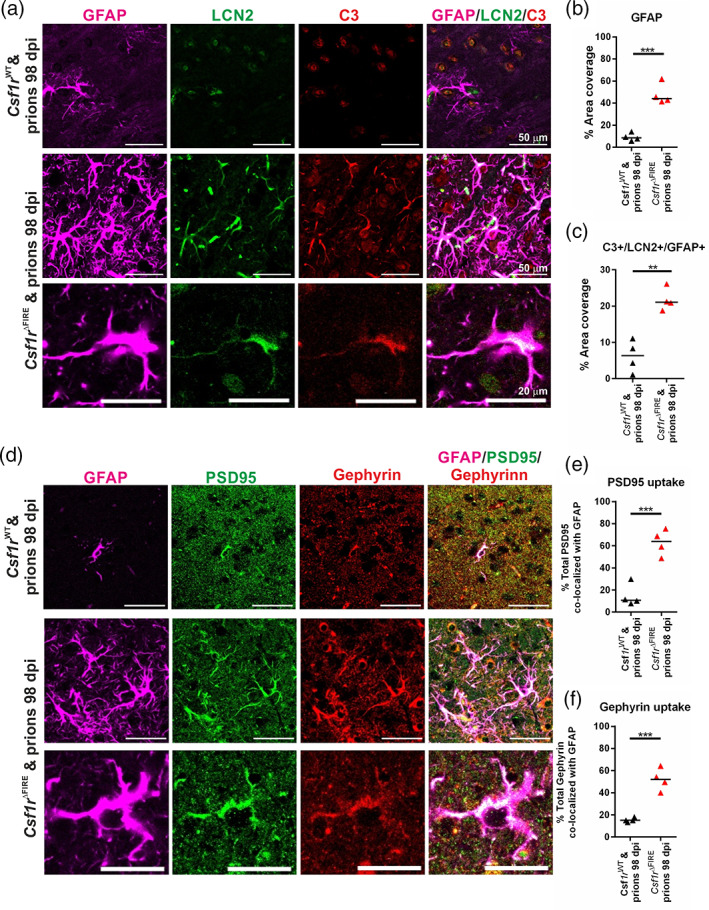
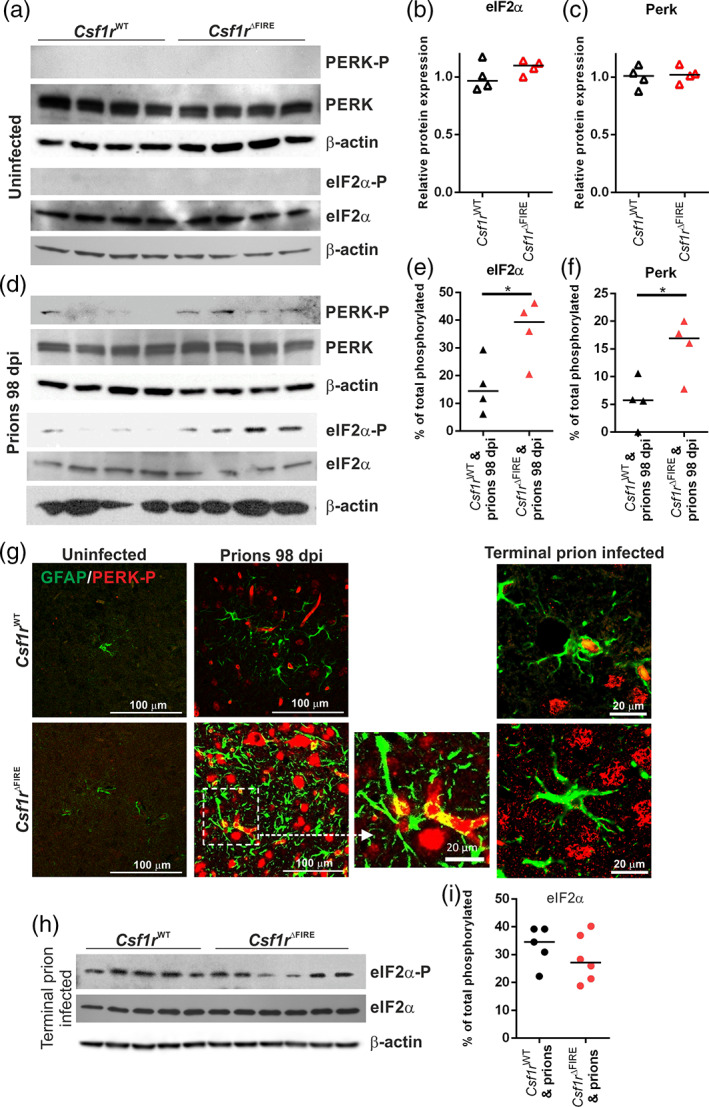
Prion diseases are transmissible, neurodegenerative disorders associated with misfolding of the prion protein. Previous studies show that reduction of microglia accelerates central nervous system (CNS) prion disease and increases the accumulation of prions in the brain, suggesting that microglia provide neuroprotection by phagocytosing and destroying prions. In Csf1rΔFIRE mice, the deletion of an enhancer within Csf1r specifically blocks microglia development, however, their brains develop normally and show none of the deficits reported in other microglia-deficient models. Csf1rΔFIRE mice were used as a refined model in which to study the impact of microglia-deficiency on CNS prion disease. Although Csf1rΔFIRE mice succumbed to CNS prion disease much earlier than wild-type mice, the accumulation of prions in their brains was reduced. Instead, astrocytes displayed earlier, non-polarized reactive activation with enhanced phagocytosis of neuronal contents and unfolded protein responses. Our data suggest that rather than simply phagocytosing and destroying prions, the microglia instead provide host-protection during CNS prion disease and restrict the harmful activities of reactive astrocytes.
Keywords: central nervous system; microglia; neurodegeneration; prion disease; reactive astrocyte.
© 2022 The Authors. GLIA published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures




References
-
- Badimon, A. , Strasburger, H. J. , Ayata, P. , Chen, X. , Nair, A. , Ikegami, A. , Hwang, P. , Chan, A. T. , Graves, S. M. , Uweru, J. O. , Ledderose, C. , Kutlu, M. G. , Wheeler, M. A. , Kahan, A. , Ishikawa, M. , Wang, Y.‐C. , Loh, Y.‐H. E. , Jiang, J. X. , Surmeier, D. J. , … Schaefer, A. (2020). Negative feedback control of neuronal activity by microglia. Nature, 586, 417–423. - PMC - PubMed
-
- Benilova, I. , Reilly, M. , Terry, C. , Wenborn, A. , Schmidt, C. , Marinho, A. T. , Risse, E. , Al‐Doujaily, H. , Wiggins De Oliveira, M. , Sandberg, M. K. , Wadsworth, J. D. F. , Jat, P. S. , & Collinge, J. (2020). Highly infectious prions are not directly neurotoxic. Proceedings of the National Academy of Sciences USA, 117(38), 23815–23822. - PMC - PubMed
-
- Beringue, V. , Demoy, M. , Lasmezas, C. I. , Gouritin, B. , Weingarten, C. , Deslys, J. P. , Andreux, J. P. , Couvreur, P. , & Dormont, D. (2000). Role of spleen macrophages in the clearance of scrapie agent early in pathogenesis. Journal of Pathology, 190(4), 495–502. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MR/M019969/1/MRC_/Medical Research Council/United Kingdom
- BB/S005471/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/D/20002173/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/D/10002071/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Miscellaneous
