

Alternative splicing of METTL3 explains apparently METTL3-independent m6A modifications in mRNA
- PMID: 35853000
- PMCID: PMC9295969
- DOI: 10.1371/journal.pbio.3001683
Alternative splicing of METTL3 explains apparently METTL3-independent m6A modifications in mRNA
Abstract
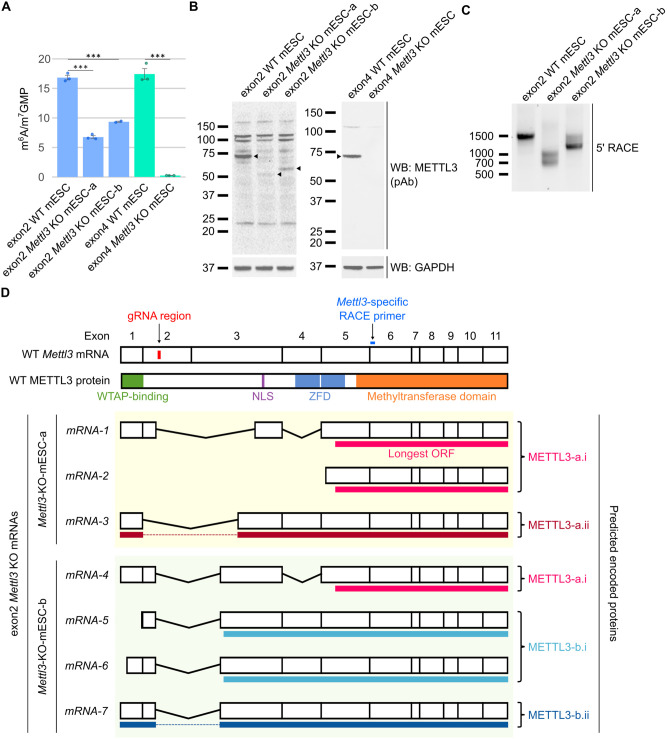
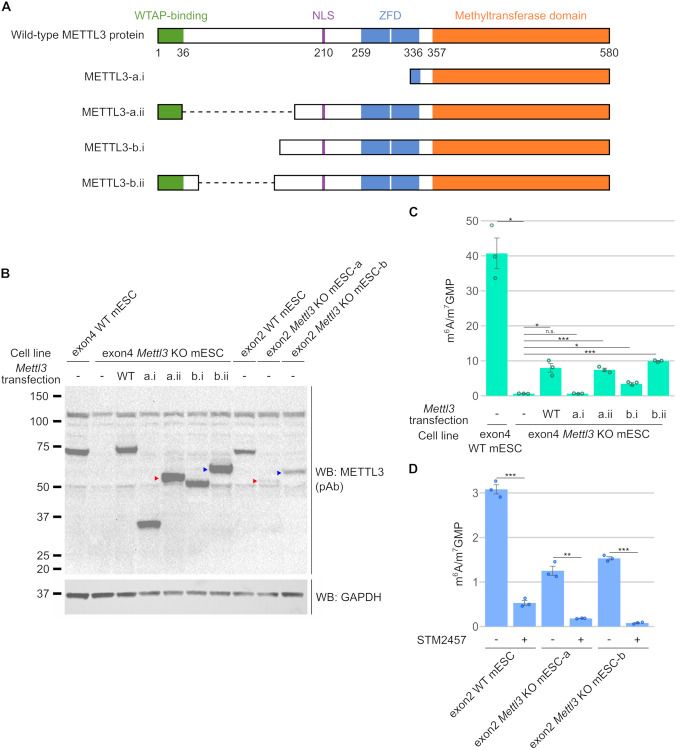
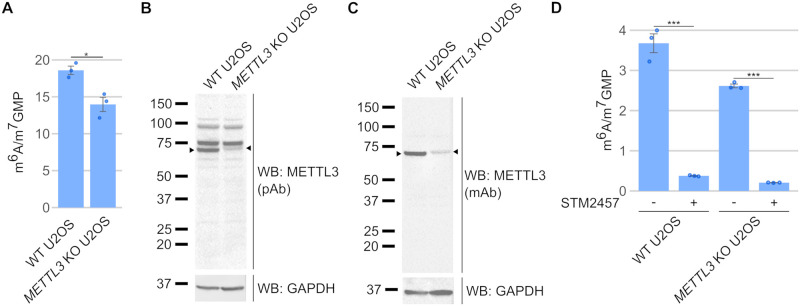
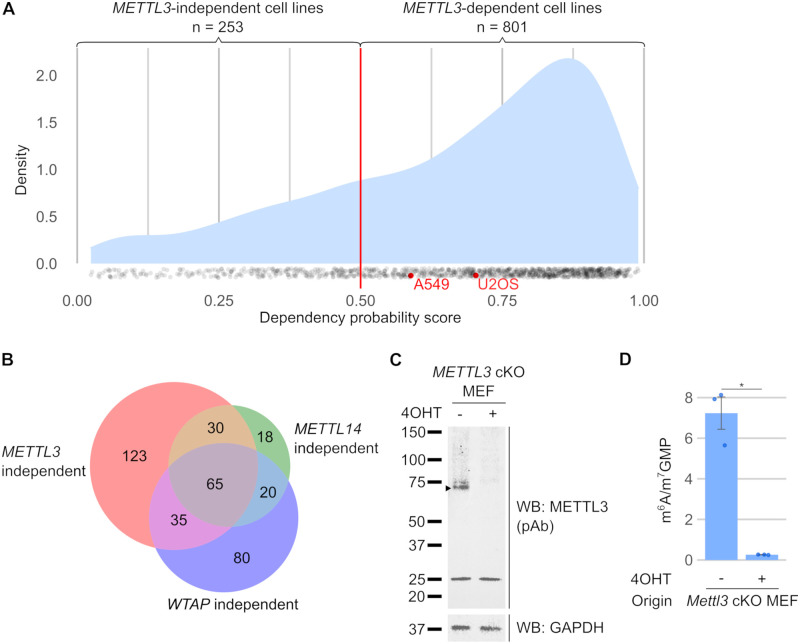
N6-methyladenosine (m6A) is a highly prevalent mRNA modification that promotes degradation of transcripts encoding proteins that have roles in cell development, differentiation, and other pathways. METTL3 is the major methyltransferase that catalyzes the formation of m6A in mRNA. As 30% to 80% of m6A can remain in mRNA after METTL3 depletion by CRISPR/Cas9-based methods, other enzymes are thought to catalyze a sizable fraction of m6A. Here, we reexamined the source of m6A in the mRNA transcriptome. We characterized mouse embryonic stem cell lines that continue to have m6A in their mRNA after Mettl3 knockout. We show that these cells express alternatively spliced Mettl3 transcript isoforms that bypass the CRISPR/Cas9 mutations and produce functionally active methyltransferases. We similarly show that other reported METTL3 knockout cell lines express altered METTL3 proteins. We find that gene dependency datasets show that most cell lines fail to proliferate after METTL3 deletion, suggesting that reported METTL3 knockout cell lines express altered METTL3 proteins rather than have full knockout. Finally, we reassessed METTL3's role in synthesizing m6A using an exon 4 deletion of Mettl3 and found that METTL3 is responsible for >95% of m6A in mRNA. Overall, these studies suggest that METTL3 is responsible for the vast majority of m6A in the transcriptome, and that remaining m6A in putative METTL3 knockout cell lines is due to the expression of altered but functional METTL3 isoforms.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: S.R.J. is scientific founder of, is advisor to, and owns equity in Gotham Therapeutics and 858 Therapeutics.
Figures
Comment in
-
Enzymes flying under the radar: Cryptic METTL3 can persist in knockout cells.PLoS Biol. 2022 Jul 20;20(7):e3001717. doi: 10.1371/journal.pbio.3001717. eCollection 2022 Jul. PLoS Biol. 2022. PMID: 35857761 Free PMC article.
References
-
- Perry RP, Kelley DE. Existence of methylated messenger RNA in mouse L cells. Cell. 1974;1:37–42. doi: 10.1016/0092-8674(74)90153-6 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
