

Genotype-phenotype correlations and disease mechanisms in PEX13-related Zellweger spectrum disorders
- PMID: 35854306
- PMCID: PMC9295491
- DOI: 10.1186/s13023-022-02415-5
Genotype-phenotype correlations and disease mechanisms in PEX13-related Zellweger spectrum disorders
Abstract
Background: Pathogenic variants in PEX-genes can affect peroxisome assembly and function and cause Zellweger spectrum disorders (ZSDs), characterized by variable phenotypes in terms of disease severity, age of onset and clinical presentations. So far, defects in at least 15 PEX-genes have been implicated in Mendelian diseases, but in some of the ultra-rare ZSD subtypes genotype-phenotype correlations and disease mechanisms remain elusive.
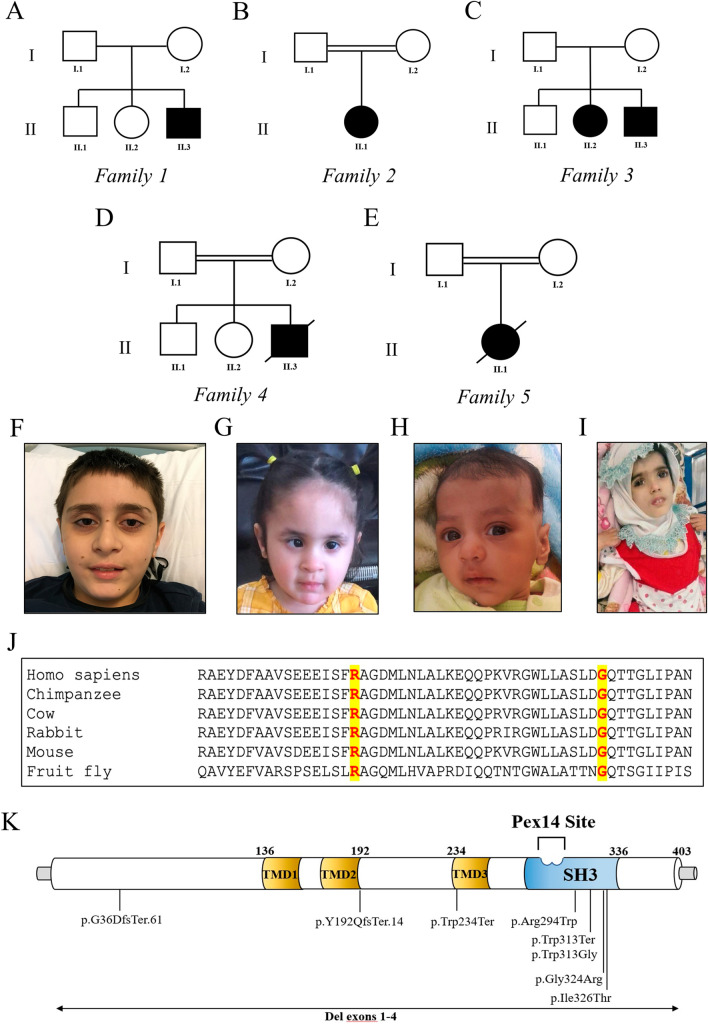
Methods: We report five families carrying biallelic variants in PEX13. The identified variants were initially evaluated by using a combination of computational approaches. Immunofluorescence and complementation studies on patient-derived fibroblasts were performed in two patients to investigate the cellular impact of the identified mutations.
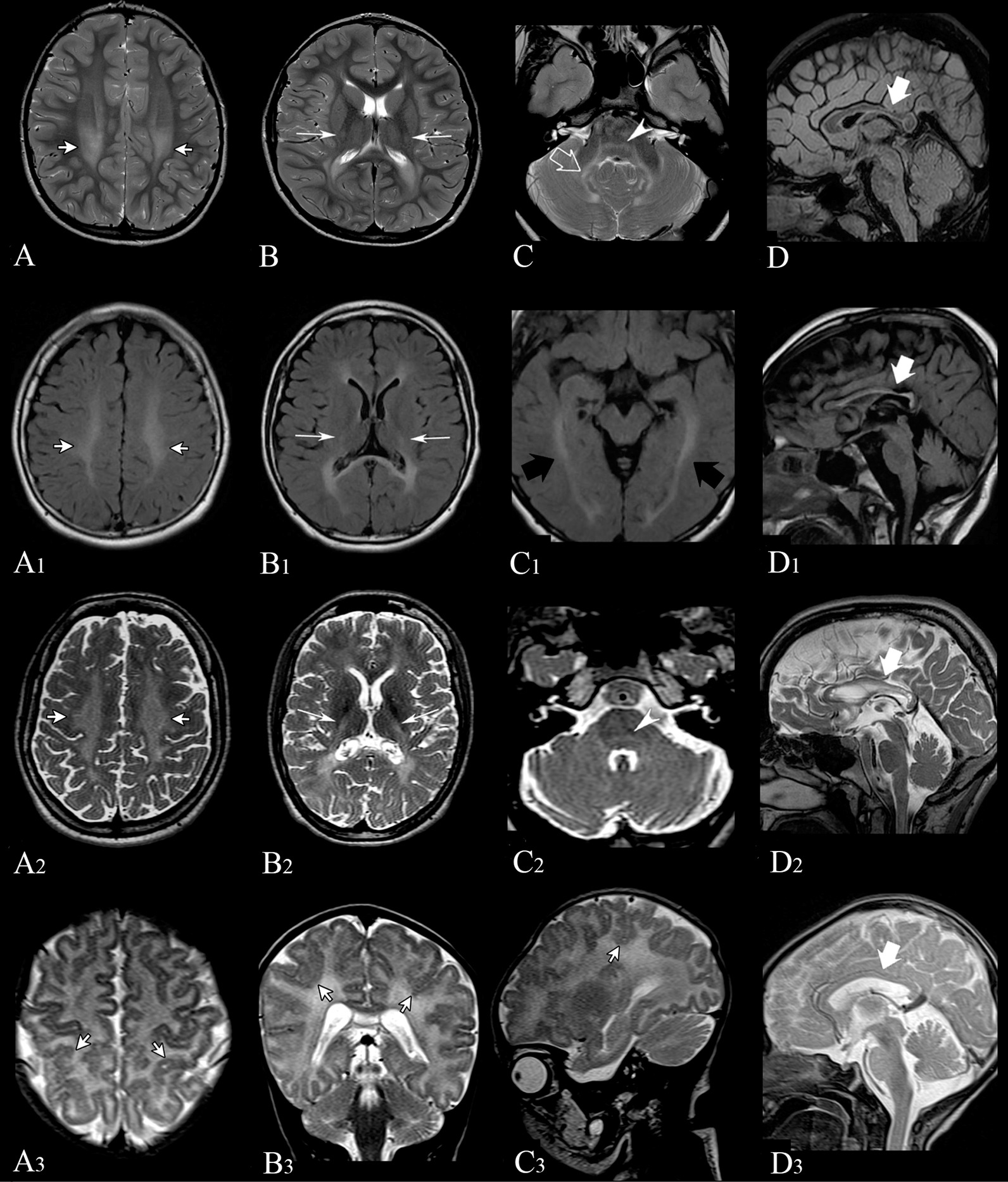
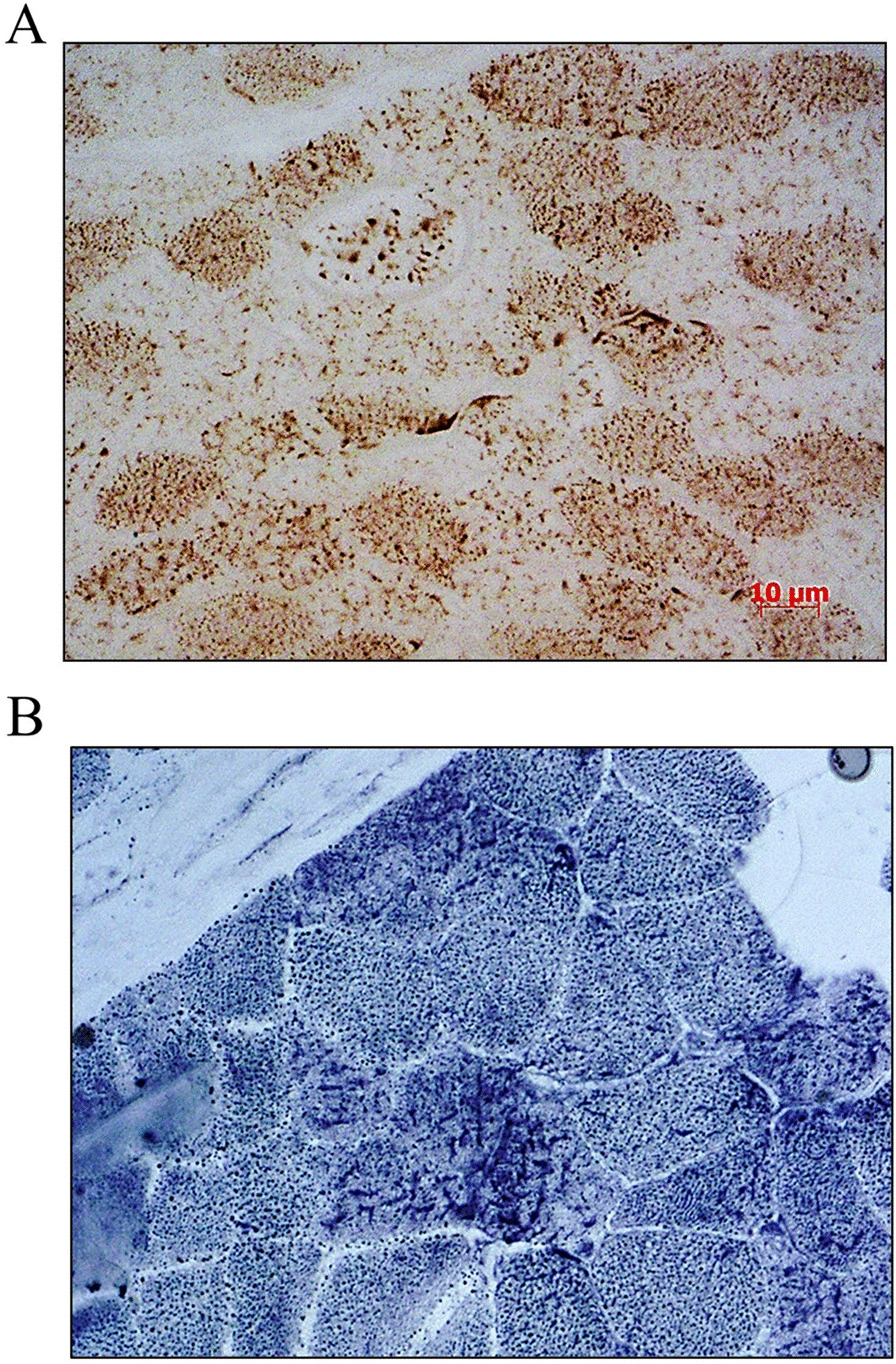
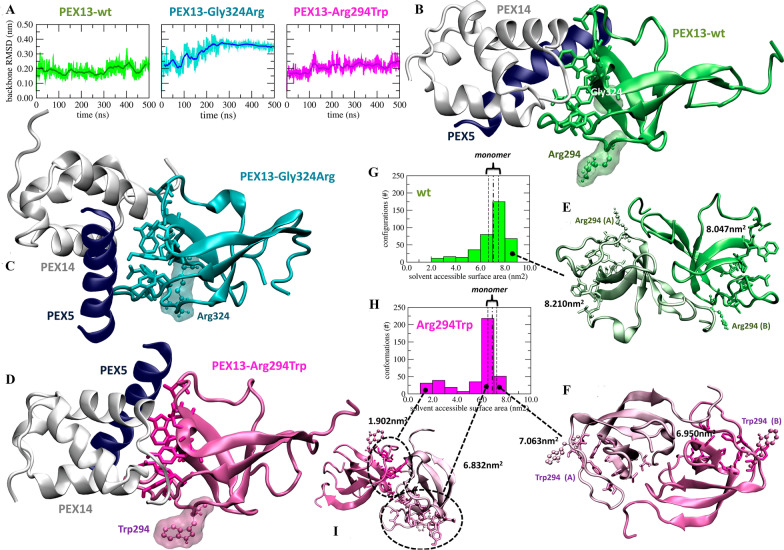
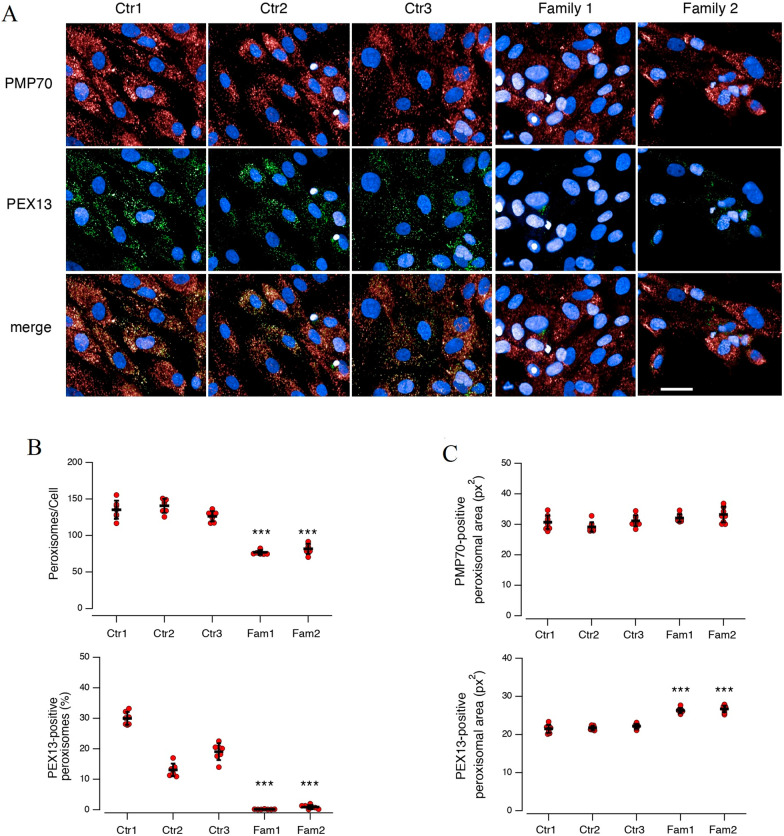
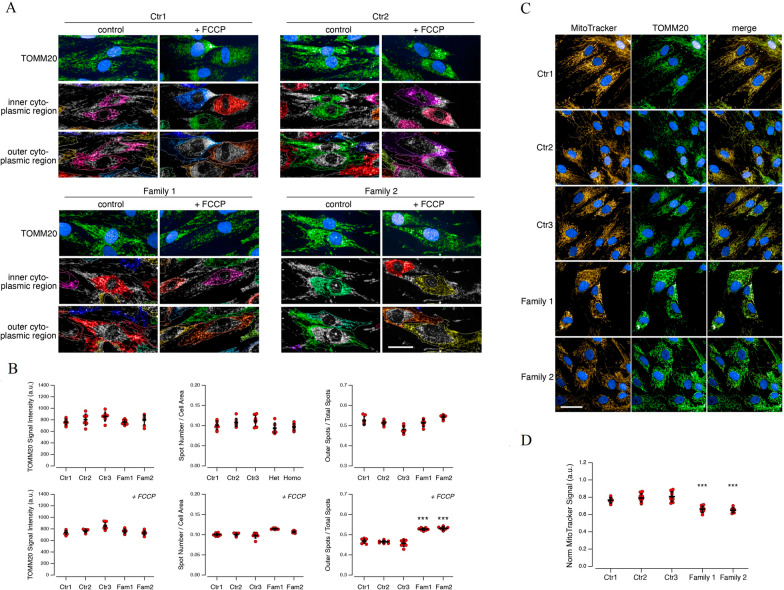
Results: Three out of five families carried a recurrent p.Arg294Trp non-synonymous variant. Individuals affected with PEX13-related ZSD presented heterogeneous clinical features, including hypotonia, developmental regression, hearing/vision impairment, progressive spasticity and brain leukodystrophy. Computational predictions highlighted the involvement of the Arg294 residue in PEX13 homodimerization, and the analysis of blind docking predicted that the p.Arg294Trp variant alters the formation of dimers, impairing the stability of the PEX13/PEX14 translocation module. Studies on muscle tissues and patient-derived fibroblasts revealed biochemical alterations of mitochondrial function and identified mislocalized mitochondria and a reduced number of peroxisomes with abnormal PEX13 concentration.
Conclusions: This study expands the phenotypic and mutational spectrum of PEX13-related ZSDs and also highlight a variety of disease mechanisms contributing to PEX13-related clinical phenotypes, including the emerging contribution of secondary mitochondrial dysfunction to the pathophysiology of ZSDs.
Keywords: PEX13; mitochondrial dysfunction; Peroxisome biogenesis disorders; Zellweger spectrum disorder.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Severe Zellweger spectrum disorder due to a novel missense variant in the PEX13 gene: A case report and the literature review.Mol Genet Genomic Med. 2024 Jan;12(1):e2315. doi: 10.1002/mgg3.2315. Epub 2023 Nov 14. Mol Genet Genomic Med. 2024. PMID: 37962062 Free PMC article.
-
Functional analysis of PEX13 mutation in a Zellweger syndrome spectrum patient reveals novel homooligomerization of PEX13 and its role in human peroxisome biogenesis.Hum Mol Genet. 2013 Oct 1;22(19):3844-57. doi: 10.1093/hmg/ddt238. Epub 2013 May 27. Hum Mol Genet. 2013. PMID: 23716570
-
Pex13 inactivation in the mouse disrupts peroxisome biogenesis and leads to a Zellweger syndrome phenotype.Mol Cell Biol. 2003 Aug;23(16):5947-57. doi: 10.1128/MCB.23.16.5947-5957.2003. Mol Cell Biol. 2003. PMID: 12897163 Free PMC article.
-
Zellweger spectrum disorders: clinical overview and management approach.Orphanet J Rare Dis. 2015 Dec 1;10:151. doi: 10.1186/s13023-015-0368-9. Orphanet J Rare Dis. 2015. PMID: 26627182 Free PMC article. Review.
-
Peroxisome biogenesis disorders in the Zellweger spectrum: An overview of current diagnosis, clinical manifestations, and treatment guidelines.Mol Genet Metab. 2016 Mar;117(3):313-21. doi: 10.1016/j.ymgme.2015.12.009. Epub 2015 Dec 23. Mol Genet Metab. 2016. PMID: 26750748 Free PMC article. Review.
Cited by
-
Spectrum of genetic alterations in patients with peroxisome biogenesis defects in the Iranian population: a case series study.BMC Med Genomics. 2025 Apr 9;18(1):67. doi: 10.1186/s12920-025-02126-3. BMC Med Genomics. 2025. PMID: 40205409 Free PMC article.
-
Variants in the WDR44 WD40-repeat domain cause a spectrum of ciliopathy by impairing ciliogenesis initiation.Nat Commun. 2024 Jan 8;15(1):365. doi: 10.1038/s41467-023-44611-2. Nat Commun. 2024. PMID: 38191484 Free PMC article.
References
-
- Steinberg SJ, Raymond GV, Braverman NE, Moser AB. Zellweger Spectrum Disorder. In: Adam MP, Ardinger HH, Pagon RA, et al., eds. GeneReviews®. University of Washington, Seattle; December 12, 2003. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
