

Broadly neutralizing antibodies target the coronavirus fusion peptide
- PMID: 35857439
- PMCID: PMC9348754
- DOI: 10.1126/science.abq3773
Broadly neutralizing antibodies target the coronavirus fusion peptide
Abstract
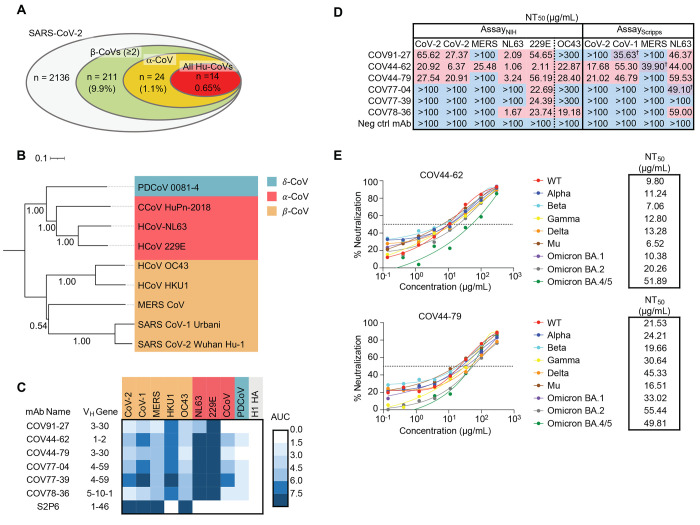
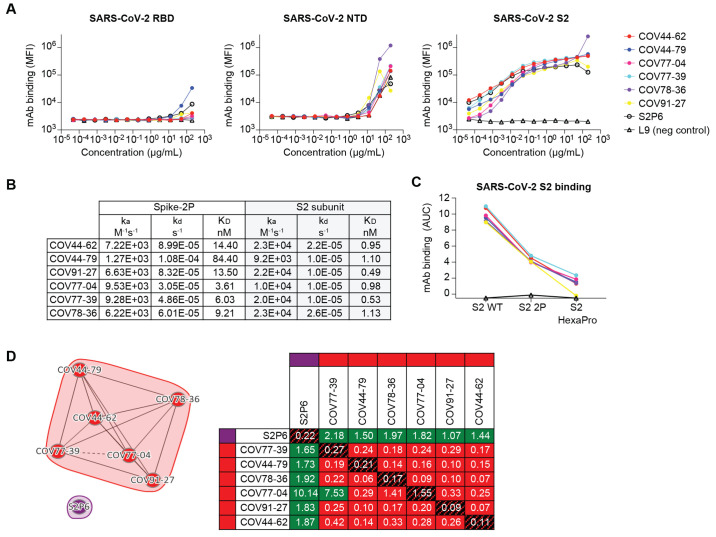
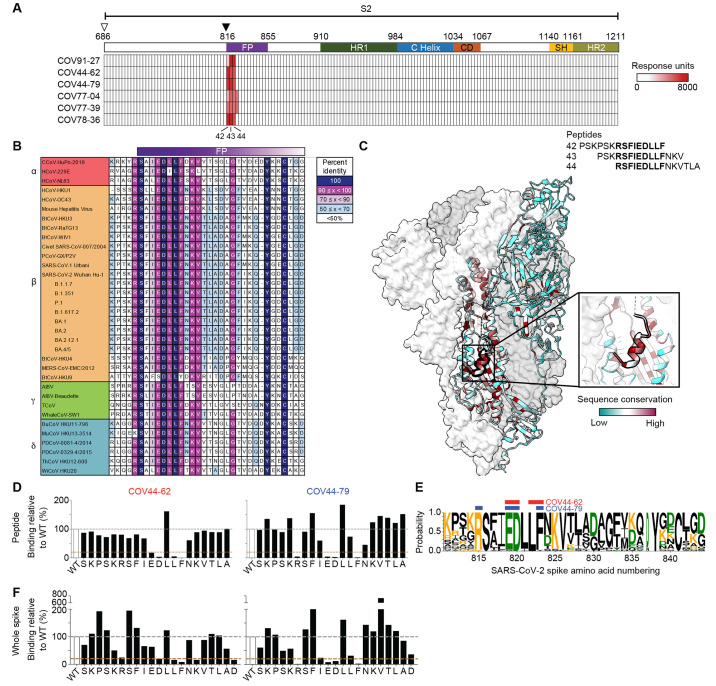
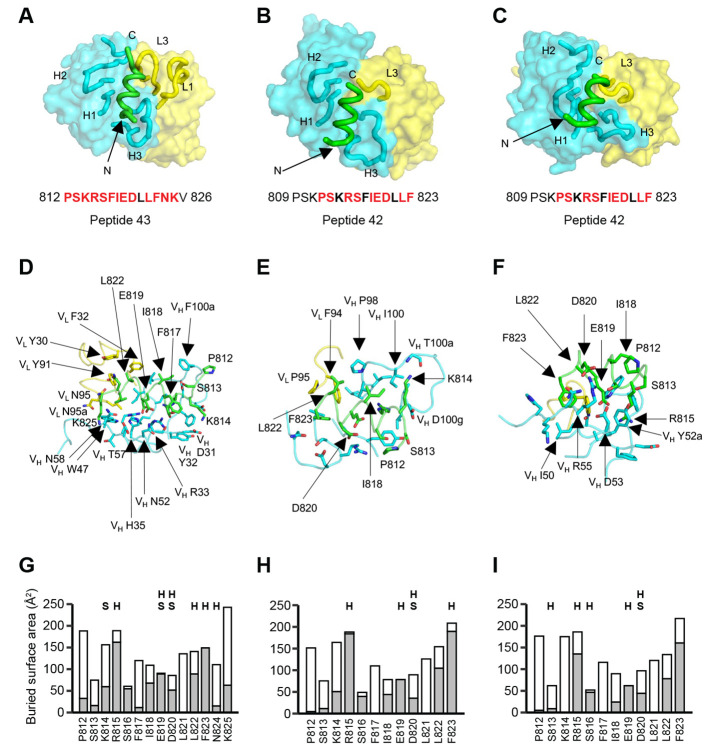
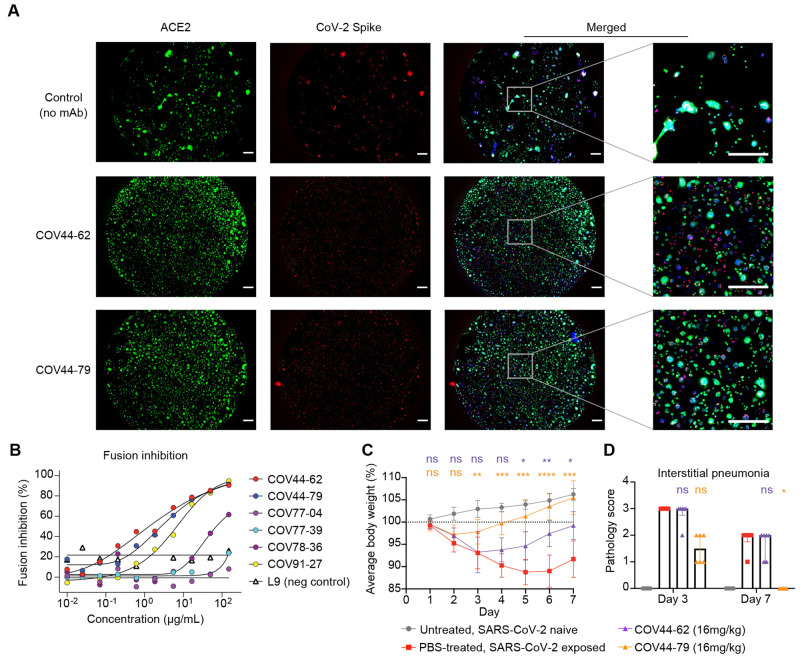
The potential for future coronavirus outbreaks highlights the need to broadly target this group of pathogens. We used an epitope-agnostic approach to identify six monoclonal antibodies that bind to spike proteins from all seven human-infecting coronaviruses. All six antibodies target the conserved fusion peptide region adjacent to the S2' cleavage site. COV44-62 and COV44-79 broadly neutralize alpha- and betacoronaviruses, including severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) Omicron subvariants BA.2 and BA.4/5, albeit with lower potency than receptor binding domain-specific antibodies. In crystal structures of COV44-62 and COV44-79 antigen-binding fragments with the SARS-CoV-2 fusion peptide, the fusion peptide epitope adopts a helical structure and includes the arginine residue at the S2' cleavage site. COV44-79 limited disease caused by SARS-CoV-2 in a Syrian hamster model. These findings highlight the fusion peptide as a candidate epitope for next-generation coronavirus vaccine development.
Figures
Update of
-
Broadly neutralizing antibodies target the coronavirus fusion peptide.bioRxiv [Preprint]. 2022 Apr 12:2022.04.11.487879. doi: 10.1101/2022.04.11.487879. bioRxiv. 2022. Update in: Science. 2022 Aug 12;377(6607):728-735. doi: 10.1126/science.abq3773. PMID: 35441178 Free PMC article. Updated. Preprint.
References
-
- Iketani S., Liu L., Guo Y., Liu L., Chan J. F.-W., Huang Y., Wang M., Luo Y., Yu J., Chu H., Chik K. K.-H., Yuen T. T.-T., Yin M. T., Sobieszczyk M. E., Huang Y., Yuen K.-Y., Wang H. H., Sheng Z., Ho D. D., Antibody evasion properties of SARS-CoV-2 Omicron sublineages. Nature 604, 553–556 (2022). 10.1038/s41586-022-04594-4 - DOI - PMC - PubMed
-
- Andrews N., Stowe J., Kirsebom F., Toffa S., Rickeard T., Gallagher E., Gower C., Kall M., Groves N., O’Connell A.-M., Simons D., Blomquist P. B., Zaidi A., Nash S., Iwani Binti Abdul Aziz N., Thelwall S., Dabrera G., Myers R., Amirthalingam G., Gharbia S., Barrett J. C., Elson R., Ladhani S. N., Ferguson N., Zambon M., Campbell C. N. J., Brown K., Hopkins S., Chand M., Ramsay M., Lopez Bernal J., Covid-19 vaccine effectiveness against the Omicron (B.1.1.529) variant. N. Engl. J. Med. 386, 1532–1546 (2022). 10.1056/NEJMoa2119451 - DOI - PMC - PubMed
-
- Takashita E., Kinoshita N., Yamayoshi S., Sakai-Tagawa Y., Fujisaki S., Ito M., Iwatsuki-Horimoto K., Chiba S., Halfmann P., Nagai H., Saito M., Adachi E., Sullivan D., Pekosz A., Watanabe S., Maeda K., Imai M., Yotsuyanagi H., Mitsuya H., Ohmagari N., Takeda M., Hasegawa H., Kawaoka Y., Efficacy of antibodies and antiviral drugs against Covid-19 Omicron variant. N. Engl. J. Med. 386, 995–998 (2022). 10.1056/NEJMc2119407 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
