

A lineage-specific Exo70 is required for receptor kinase-mediated immunity in barley
- PMID: 35857460
- PMCID: PMC9258809
- DOI: 10.1126/sciadv.abn7258
A lineage-specific Exo70 is required for receptor kinase-mediated immunity in barley
Abstract
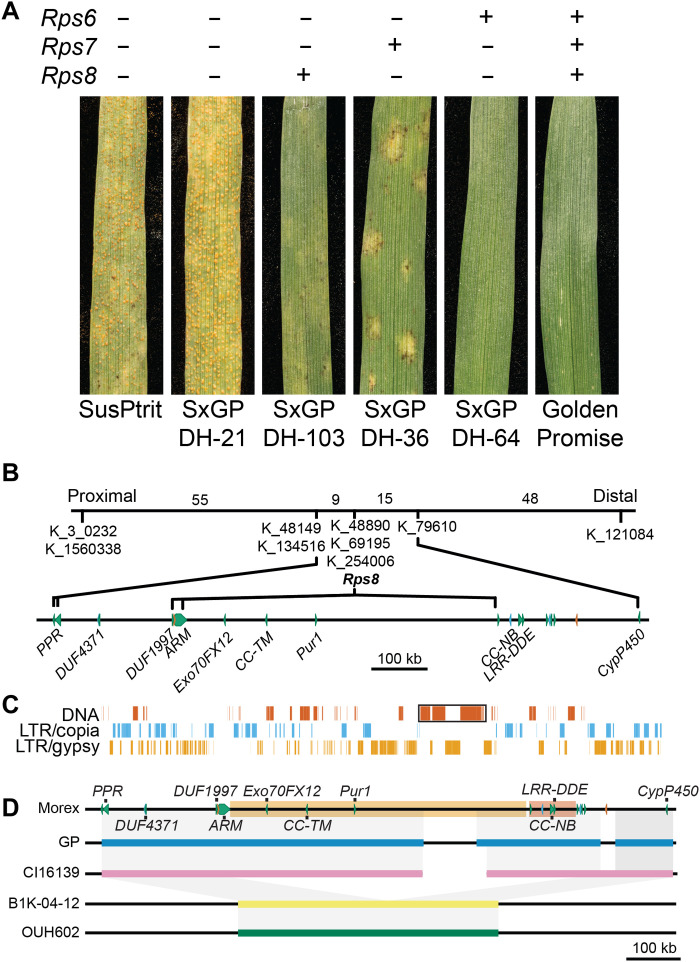
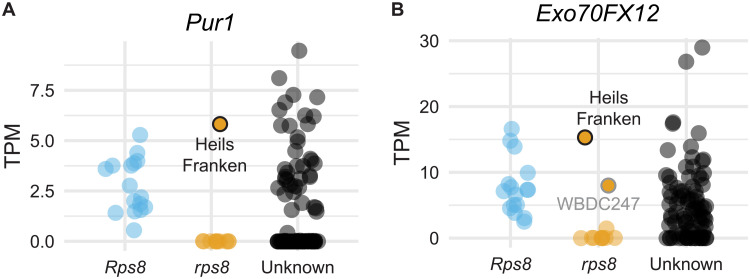
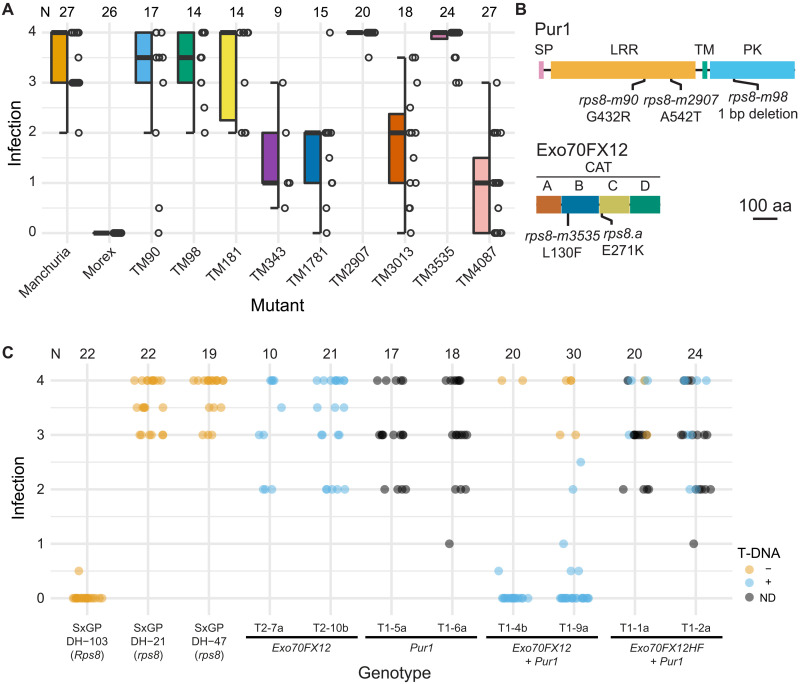
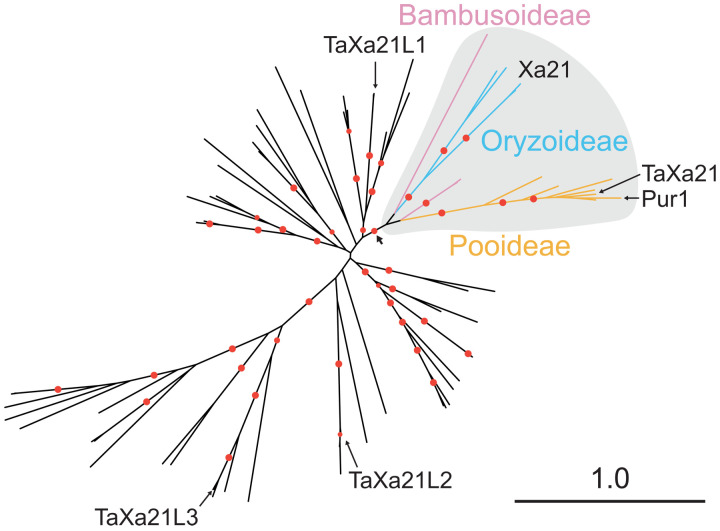
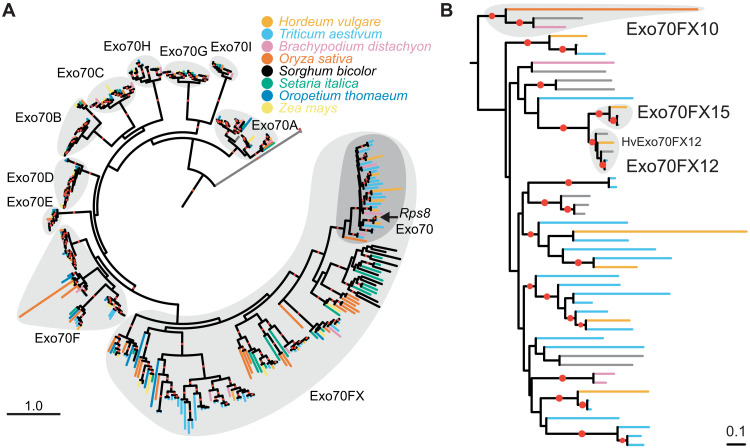
In the evolution of land plants, the plant immune system has experienced expansion in immune receptor and signaling pathways. Lineage-specific expansions have been observed in diverse gene families that are potentially involved in immunity but lack causal association. Here, we show that Rps8-mediated resistance in barley to the pathogen Puccinia striiformis f. sp. tritici (wheat stripe rust) is conferred by a genetic module: Pur1 and Exo70FX12, which are together necessary and sufficient. Pur1 encodes a leucine-rich repeat receptor kinase and is the ortholog of rice Xa21, and Exo70FX12 belongs to the Poales-specific Exo70FX clade. The Exo70FX clade emerged after the divergence of the Bromeliaceae and Poaceae and comprises from 2 to 75 members in sequenced grasses. These results demonstrate the requirement of a lineage-specific Exo70FX12 in Pur1-mediated immunity and suggest that the Exo70FX clade may have evolved a specialized role in receptor kinase signaling.
Figures
References
-
- Dodds P. N., Rathjen J. P., Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 11, 539–548 (2010). - PubMed
-
- Lo Presti L., Lanver D., Schweizer G., Tanaka S., Liang L., Tollot M., Zuccaro A., Reissmann S., Kahmann R., Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 66, 513–545 (2015). - PubMed
-
- Cui H., Tsuda K., Parker J. E., Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 66, 487–511 (2015). - PubMed
-
- Zipfel C., Oldroyd G. E., Plant signalling in symbiosis and immunity. Nature 543, 328–336 (2017). - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
