

Cis-regulatory chromatin loops analysis identifies GRHL3 as a master regulator of surface epithelium commitment
- PMID: 35857527
- PMCID: PMC9278850
- DOI: 10.1126/sciadv.abo5668
Cis-regulatory chromatin loops analysis identifies GRHL3 as a master regulator of surface epithelium commitment
Abstract
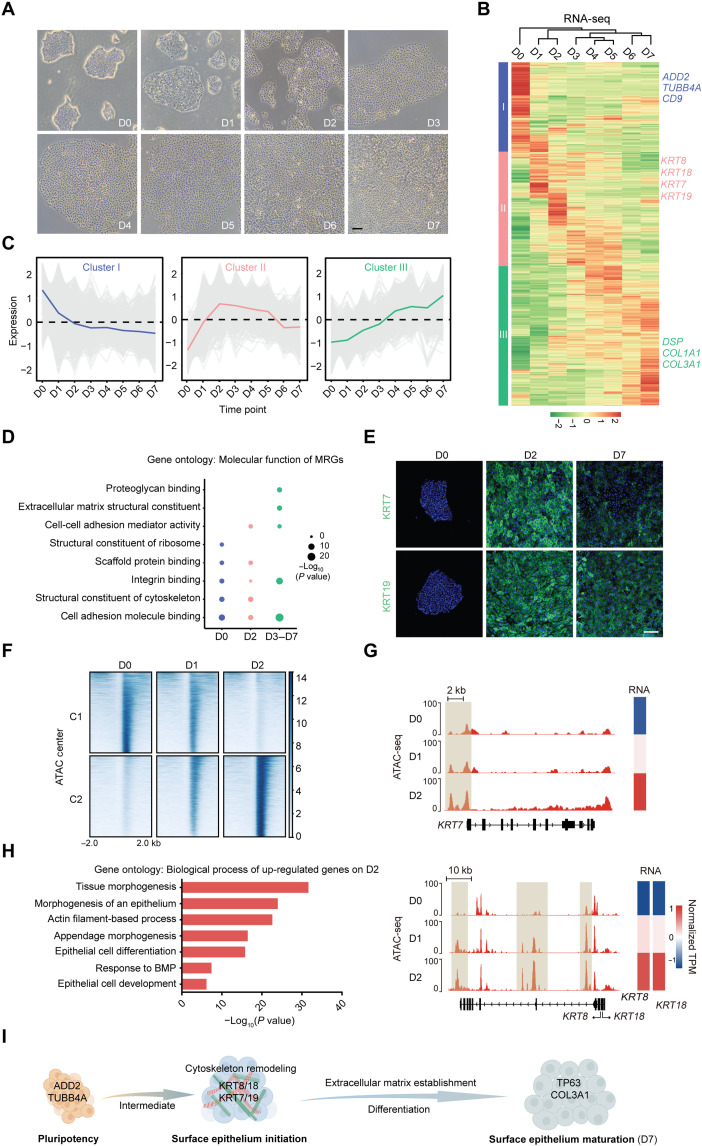
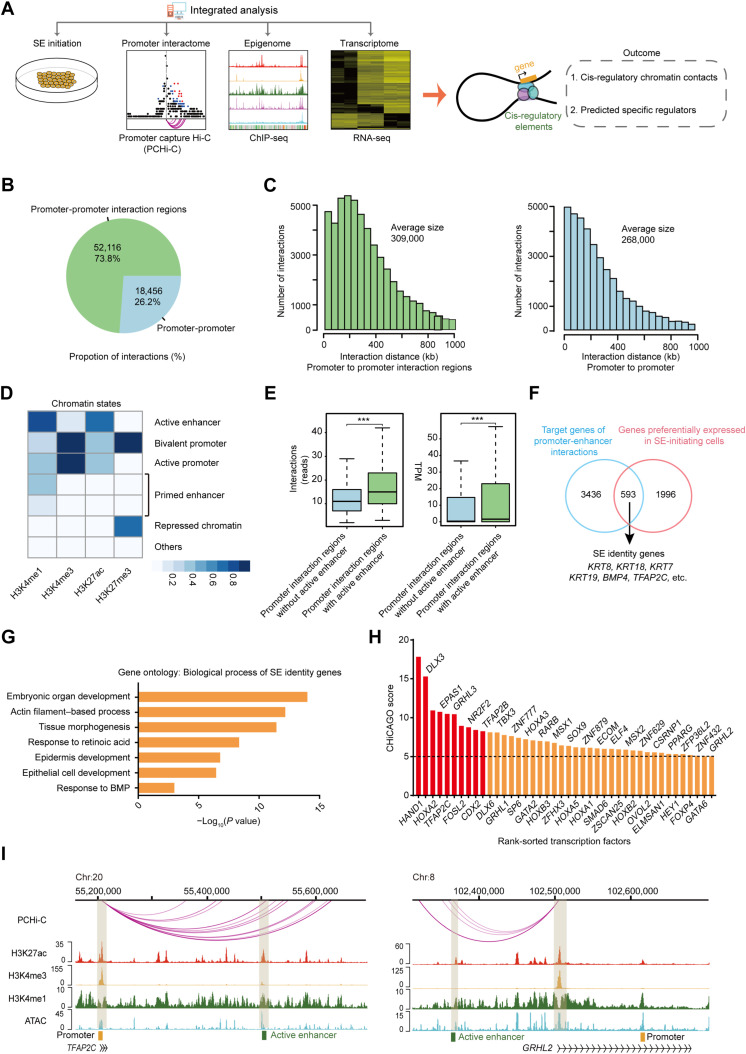
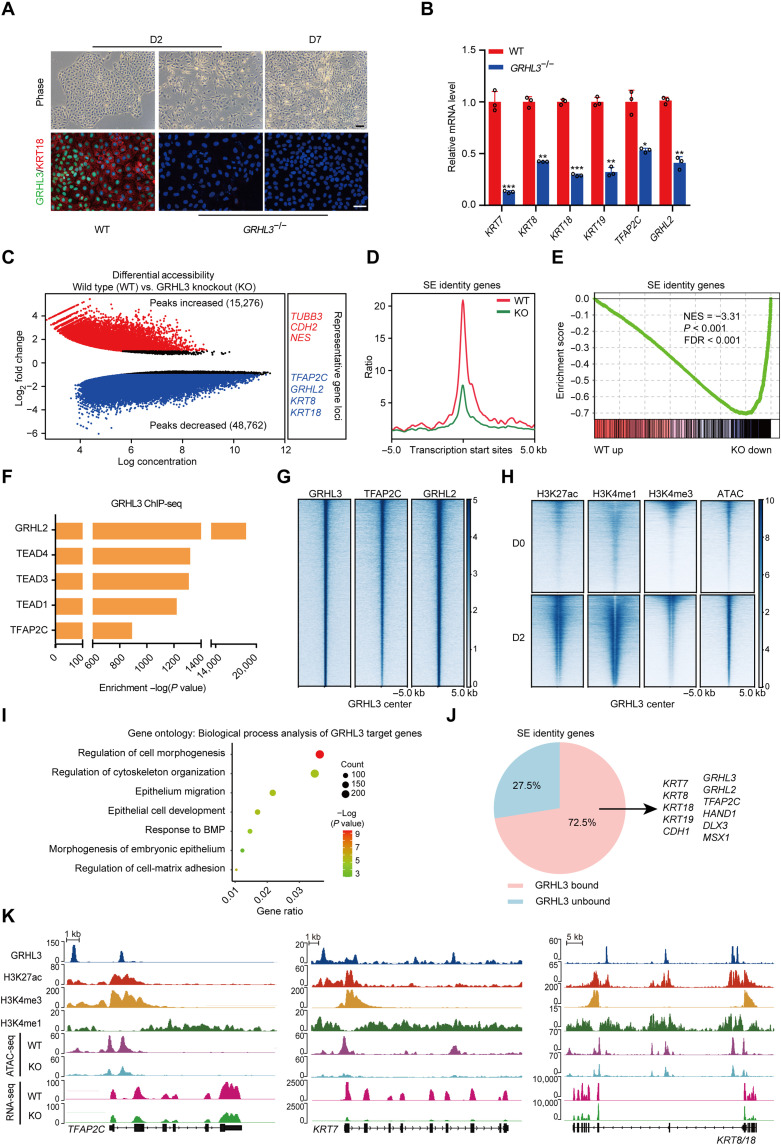
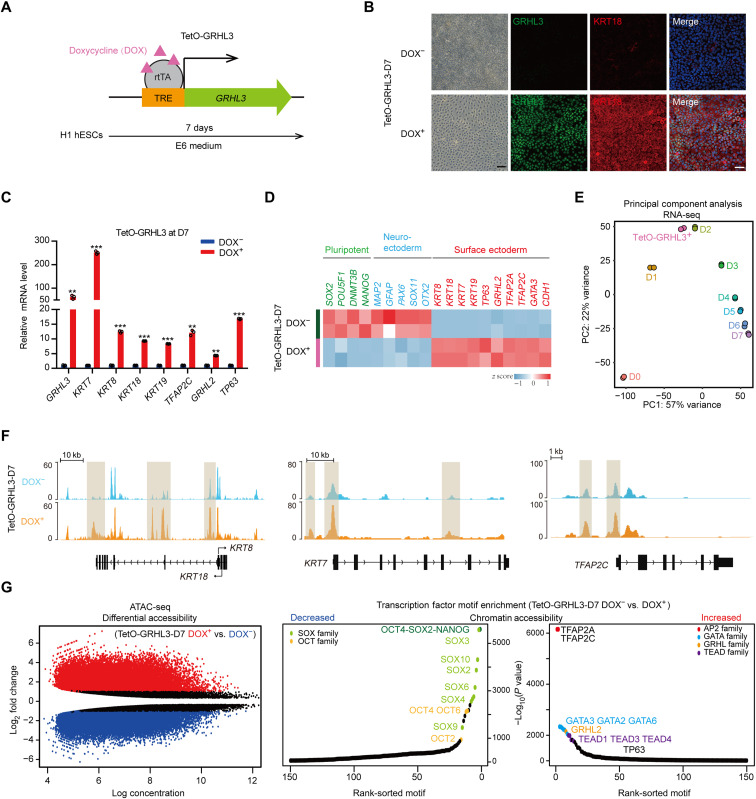
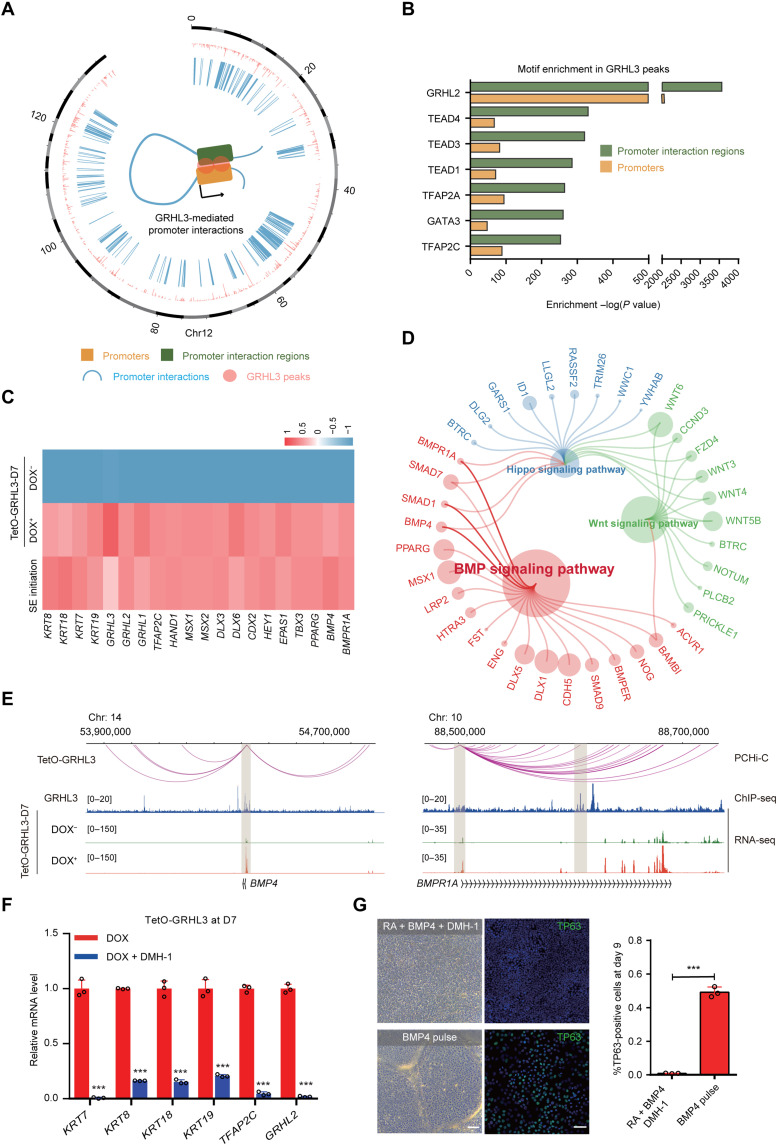
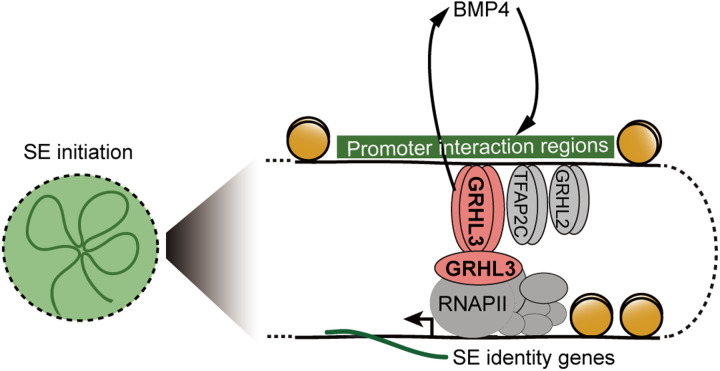
Understanding the regulatory network of cell fate acquisition remains a major challenge. Using the induction of surface epithelium (SE) from human embryonic stem cells as a paradigm, we show that the dynamic changes in morphology-related genes (MRGs) closely correspond to SE fate transitions. The marked remodeling of cytoskeleton indicates the initiation of SE differentiation. By integrating promoter interactions, epigenomic features, and transcriptome, we delineate an SE-specific cis-regulatory network and identify grainyhead-like 3 (GRHL3) as an initiation factor sufficient to drive SE commitment. Mechanically, GRHL3 primes the SE chromatin accessibility landscape and activates SE-initiating gene expression. In addition, the evaluation of GRHL3-mediated promoter interactions unveils a positive feedback loop of GRHL3 and bone morphogenetic protein 4 on SE fate decisions. Our work proposes a concept that MRGs could be used to identify cell fate transitions and provides insights into regulatory principles of SE lineage development and stem cell-based regenerative medicine.
Figures
References
-
- Li L., Wang Y., Torkelson J. L., Shankar G., Pattison J. M., Zhen H. H., Fang F., Duren Z., Xin J., Gaddam S., Melo S. P., Piekos S. N., Li J., Liaw E. J., Chen L., Li R., Wernig M., Wong W. H., Chang H. Y., Oro A. E., TFAP2C- and p63-dependent networks sequentially rearrange chromatin landscapes to drive human epidermal lineage commitment. Cell Stem Cell 24, 271–284.e8 (2019). - PMC - PubMed
-
- Fan X., Wang D., Burgmaier J. E., Teng Y., Romano R. A., Sinha S., Yi R., Single cell and open chromatin analysis reveals molecular origin of epidermal cells of the skin. Dev. Cell 47, 133 (2018). - PubMed
-
- Schoenfelder S., Furlan-Magaril M., Mifsud B., Tavares-Cadete F., Sugar R., Javierre B. M., Nagano T., Katsman Y., Sakthidevi M., Wingett S. W., Dimitrova E., Dimond A., Edelman L. B., Elderkin S., Tabbada K., Darbo E., Andrews S., Herman B., Higgs A., LeProust E., Osborne C. S., Mitchell J. A., Luscombe N. M., Fraser P., The pluripotent regulatory circuitry connecting promoters to their long-range interacting elements. Genome Res. 25, 582–597 (2015). - PMC - PubMed
-
- Rubin A. J., Barajas B. C., Furlan-Magaril M., Lopez-Pajares V., Mumbach M. R., Howard I., Kim D. S., Boxer L. D., Cairns J., Spivakov M., Wingett S. W., Shi M., Zhao Z., Greenleaf W. J., Kundaje A., Snyder M., Chang H. Y., Fraser P., Khavari P. A., Lineage-specific dynamic and pre-established enhancer-promoter contacts cooperate in terminal differentiation. Nat. Genet. 49, 1522–1528 (2017). - PMC - PubMed
-
- Freire-Pritchett P., Schoenfelder S., Varnai C., Wingett S. W., Cairns J., Collier A. J., Garcia-Vilchez R., Furlan-Magaril M., Osborne C. S., Fraser P., Rugg-Gunn P. J., Spivakov M., Global reorganisation of cis-regulatory units upon lineage commitment of human embryonic stem cells. eLife 6, e21926 (2017). - PMC - PubMed
LinkOut - more resources
Full Text Sources
