

Structural analyses of human ryanodine receptor type 2 channels reveal the mechanisms for sudden cardiac death and treatment
- PMID: 35857850
- PMCID: PMC9299551
- DOI: 10.1126/sciadv.abo1272
Structural analyses of human ryanodine receptor type 2 channels reveal the mechanisms for sudden cardiac death and treatment
Abstract
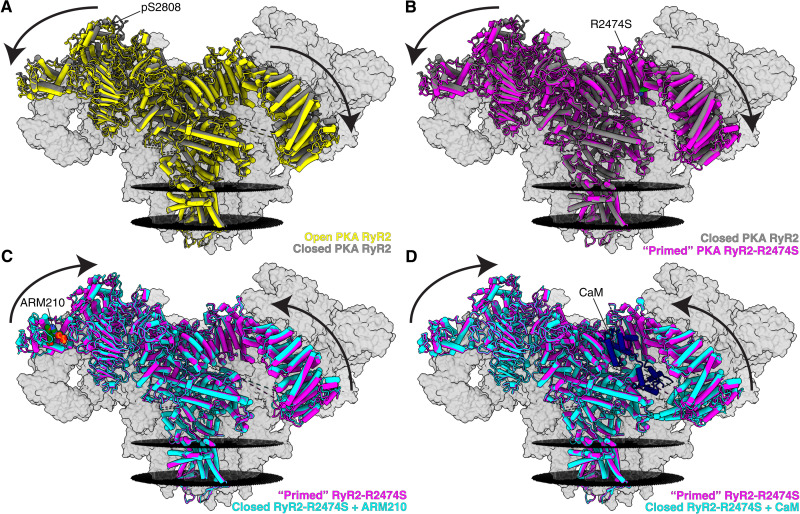
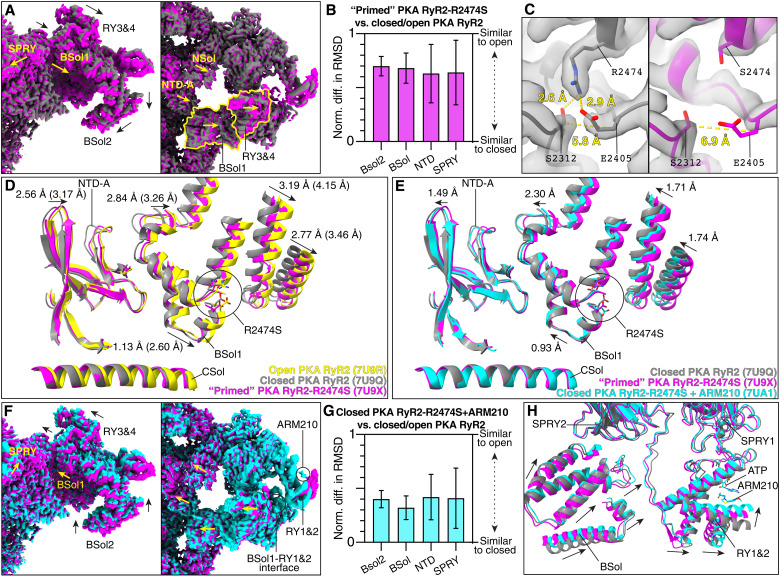
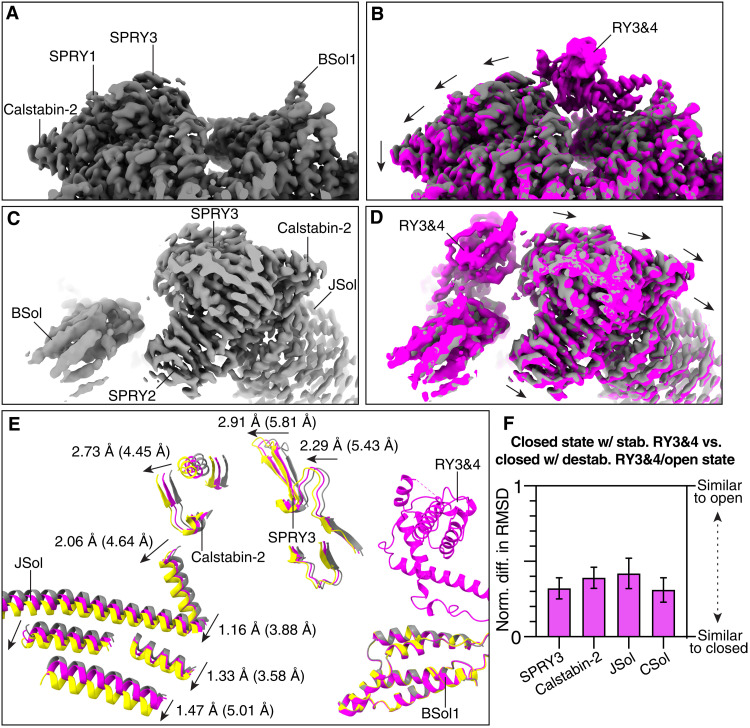
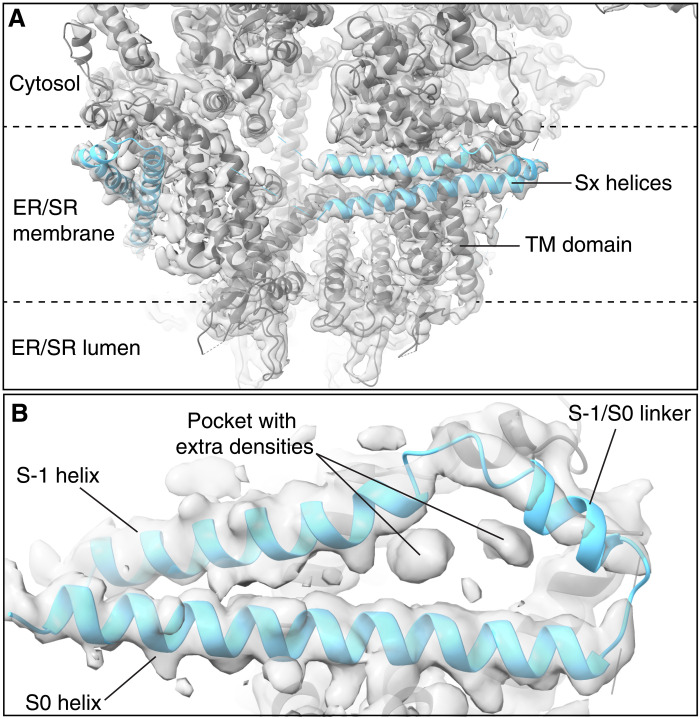
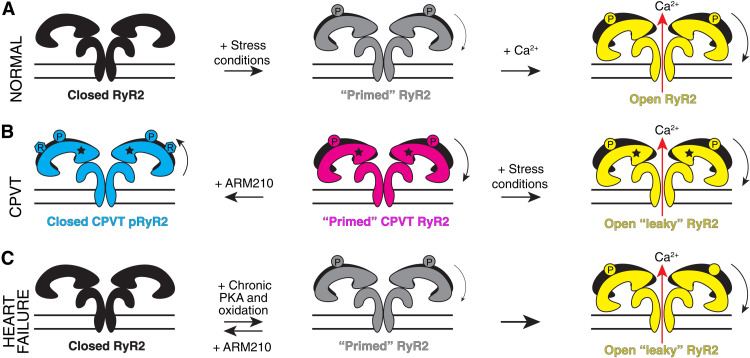
Ryanodine receptor type 2 (RyR2) mutations have been linked to an inherited form of exercise-induced sudden cardiac death called catecholaminergic polymorphic ventricular tachycardia (CPVT). CPVT results from stress-induced sarcoplasmic reticular Ca2+ leak via the mutant RyR2 channels during diastole. We present atomic models of human wild-type (WT) RyR2 and the CPVT mutant RyR2-R2474S determined by cryo-electron microscopy with overall resolutions in the range of 2.6 to 3.6 Å, and reaching local resolutions of 2.25 Å, unprecedented for RyR2 channels. Under nonactivating conditions, the RyR2-R2474S channel is in a "primed" state between the closed and open states of WT RyR2, rendering it more sensitive to activation that results in stress-induced Ca2+ leak. The Rycal drug ARM210 binds to RyR2-R2474S, reverting the primed state toward the closed state. Together, these studies provide a mechanism for CPVT and for the therapeutic actions of ARM210.
Figures

Similar articles
-
Calcium leak through ryanodine receptors leads to atrial fibrillation in 3 mouse models of catecholaminergic polymorphic ventricular tachycardia.Circ Res. 2012 Aug 31;111(6):708-17. doi: 10.1161/CIRCRESAHA.112.273342. Epub 2012 Jul 24. Circ Res. 2012. PMID: 22828895 Free PMC article.
-
Catecholaminergic polymorphic ventricular tachycardia-linked ryanodine receptor variants exhibit domain-specific calcium leak and calmodulin affinity properties.J Physiol. 2025 May 5. doi: 10.1113/JP288499. Online ahead of print. J Physiol. 2025. PMID: 40320903
-
Leaky Ca2+ release channel/ryanodine receptor 2 causes seizures and sudden cardiac death in mice.J Clin Invest. 2008 Jun;118(6):2230-45. doi: 10.1172/JCI35346. J Clin Invest. 2008. PMID: 18483626 Free PMC article.
-
Catecholaminergic polymorphic ventricular tachycardia: recent mechanistic insights.Cardiovasc Res. 2005 Aug 15;67(3):379-87. doi: 10.1016/j.cardiores.2005.04.027. Cardiovasc Res. 2005. PMID: 15913575 Review.
-
Involvement of the cardiac ryanodine receptor/calcium release channel in catecholaminergic polymorphic ventricular tachycardia.J Cell Physiol. 2002 Jan;190(1):1-6. doi: 10.1002/jcp.10031. J Cell Physiol. 2002. PMID: 11807805 Review.
Cited by
-
Arrhythmogenic mechanism of a novel ryanodine receptor mutation underlying sudden cardiac death.Europace. 2023 Jul 4;25(7):euad220. doi: 10.1093/europace/euad220. Europace. 2023. PMID: 37466361 Free PMC article.
-
Structural identification of the RY12 domain of RyR1 as an ADP sensor and the target of the malignant hyperthermia therapeutic dantrolene.bioRxiv [Preprint]. 2024 Oct 24:2024.10.21.619409. doi: 10.1101/2024.10.21.619409. bioRxiv. 2024. PMID: 39484412 Free PMC article. Preprint.
-
Location, location, and activation of a channel by calcium.Proc Natl Acad Sci U S A. 2022 Oct 25;119(43):e2214826119. doi: 10.1073/pnas.2214826119. Epub 2022 Oct 10. Proc Natl Acad Sci U S A. 2022. PMID: 36215521 Free PMC article. No abstract available.
-
A dual-targeted drug inhibits cardiac ryanodine receptor Ca2+ leak but activates SERCA2a Ca2+ uptake.Life Sci Alliance. 2023 Nov 27;7(2):e202302278. doi: 10.26508/lsa.202302278. Print 2024 Feb. Life Sci Alliance. 2023. PMID: 38012000 Free PMC article.
-
Personalized medicine in the dish to prevent calcium leak associated with short-coupled polymorphic ventricular tachycardia in patient-derived cardiomyocytes.Stem Cell Res Ther. 2023 Sep 23;14(1):266. doi: 10.1186/s13287-023-03502-5. Stem Cell Res Ther. 2023. PMID: 37740238 Free PMC article.
References
-
- Otsu K., Willard H. F., Khanna V. K., Zorzato F., Green N. M., MacLennan D. H., Molecular cloning of cDNA encoding the Ca2+ release channel (ryanodine receptor) of rabbit cardiac muscle sarcoplasmic reticulum. J. Biol. Chem. 265, 13472–13483 (1990). - PubMed
-
- Nakai J., Imagawa T., Hakamat Y., Shigekawa M., Takeshima H., Numa S., Primary structure and functional expression from cDNA of the cardiac ryanodine receptor/calcium release channel. FEBS Lett. 271, 169–177 (1990). - PubMed
-
- Fabiato A., Calcium-induced release of calcium from the cardiac sarcoplasmic reticulum. Am. J. Physiol. 245, C1–C14 (1983). - PubMed
-
- Marx S. O., Reiken S., Hisamatsu Y., Jayaraman T., Burkhoff D., Rosemblit N., Marks A. R., PKA phosphorylation dissociates FKBP12.6 from the calcium release channel (ryanodine receptor): Defective regulation in failing hearts. Cell 101, 365–376 (2000). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
