

Pareto optimality, economy-effectiveness trade-offs and ion channel degeneracy: improving population modelling for single neurons
- PMID: 35857898
- PMCID: PMC9277232
- DOI: 10.1098/rsob.220073
Pareto optimality, economy-effectiveness trade-offs and ion channel degeneracy: improving population modelling for single neurons
Abstract
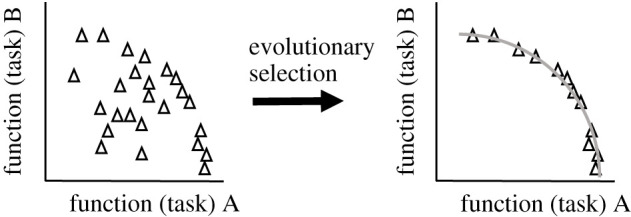
Neurons encounter unavoidable evolutionary trade-offs between multiple tasks. They must consume as little energy as possible while effectively fulfilling their functions. Cells displaying the best performance for such multi-task trade-offs are said to be Pareto optimal, with their ion channel configurations underpinning their functionality. Ion channel degeneracy, however, implies that multiple ion channel configurations can lead to functionally similar behaviour. Therefore, instead of a single model, neuroscientists often use populations of models with distinct combinations of ionic conductances. This approach is called population (database or ensemble) modelling. It remains unclear, which ion channel parameters in the vast population of functional models are more likely to be found in the brain. Here we argue that Pareto optimality can serve as a guiding principle for addressing this issue by helping to identify the subpopulations of conductance-based models that perform best for the trade-off between economy and functionality. In this way, the high-dimensional parameter space of neuronal models might be reduced to geometrically simple low-dimensional manifolds, potentially explaining experimentally observed ion channel correlations. Conversely, Pareto inference might also help deduce neuronal functions from high-dimensional Patch-seq data. In summary, Pareto optimality is a promising framework for improving population modelling of neurons and their circuits.
Keywords: Pareto front; energy efficiency; ion channel correlations; multi-objective optimization; parameter space; performance space.
Conflict of interest statement
We declare we have no competing interests.
Figures







Similar articles
-
Evolutionary trade-offs and the structure of polymorphisms.Philos Trans R Soc Lond B Biol Sci. 2018 May 26;373(1747):20170105. doi: 10.1098/rstb.2017.0105. Philos Trans R Soc Lond B Biol Sci. 2018. PMID: 29632259 Free PMC article.
-
The geometry of the Pareto front in biological phenotype space.Ecol Evol. 2013 Jun;3(6):1471-83. doi: 10.1002/ece3.528. Epub 2013 Apr 17. Ecol Evol. 2013. PMID: 23789060 Free PMC article.
-
Evolutionary trade-offs, Pareto optimality, and the geometry of phenotype space.Science. 2012 Jun 1;336(6085):1157-60. doi: 10.1126/science.1217405. Epub 2012 Apr 26. Science. 2012. PMID: 22539553
-
Neural optimization: Understanding trade-offs with Pareto theory.Curr Opin Neurobiol. 2021 Dec;71:84-91. doi: 10.1016/j.conb.2021.08.008. Epub 2021 Oct 21. Curr Opin Neurobiol. 2021. PMID: 34688051 Review.
-
Searching for the Pareto frontier in multi-objective protein design.Biophys Rev. 2017 Aug;9(4):339-344. doi: 10.1007/s12551-017-0288-0. Epub 2017 Aug 10. Biophys Rev. 2017. PMID: 28799089 Free PMC article. Review.
Cited by
-
The emergence of nonlinear evolutionary trade-offs and the maintenance of genetic polymorphisms.Biol Lett. 2024 Dec;20(12):20240296. doi: 10.1098/rsbl.2024.0296. Epub 2024 Dec 4. Biol Lett. 2024. PMID: 39626761
-
Biological complexity facilitates tuning of the neuronal parameter space.PLoS Comput Biol. 2023 Jul 3;19(7):e1011212. doi: 10.1371/journal.pcbi.1011212. eCollection 2023 Jul. PLoS Comput Biol. 2023. PMID: 37399220 Free PMC article.
-
Homeostatic regulation of neuronal function: importance of degeneracy and pleiotropy.Front Cell Neurosci. 2023 Jun 2;17:1184563. doi: 10.3389/fncel.2023.1184563. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37333893 Free PMC article. Review.
-
The Emergence of Non-Linear Evolutionary Trade-offs and the Maintenance of Genetic Polymorphisms.bioRxiv [Preprint]. 2024 Jun 1:2024.05.29.595890. doi: 10.1101/2024.05.29.595890. bioRxiv. 2024. Update in: Biol Lett. 2024 Dec;20(12):20240296. doi: 10.1098/rsbl.2024.0296. PMID: 38853830 Free PMC article. Updated. Preprint.
-
Heterogeneous off-target impact of ion-channel deletion on intrinsic properties of hippocampal model neurons that self-regulate calcium.Front Cell Neurosci. 2023 Oct 10;17:1241450. doi: 10.3389/fncel.2023.1241450. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37904732 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
