

Sox9 directs divergent epigenomic states in brain tumor subtypes
- PMID: 35858326
- PMCID: PMC9303974
- DOI: 10.1073/pnas.2202015119
Sox9 directs divergent epigenomic states in brain tumor subtypes
Abstract
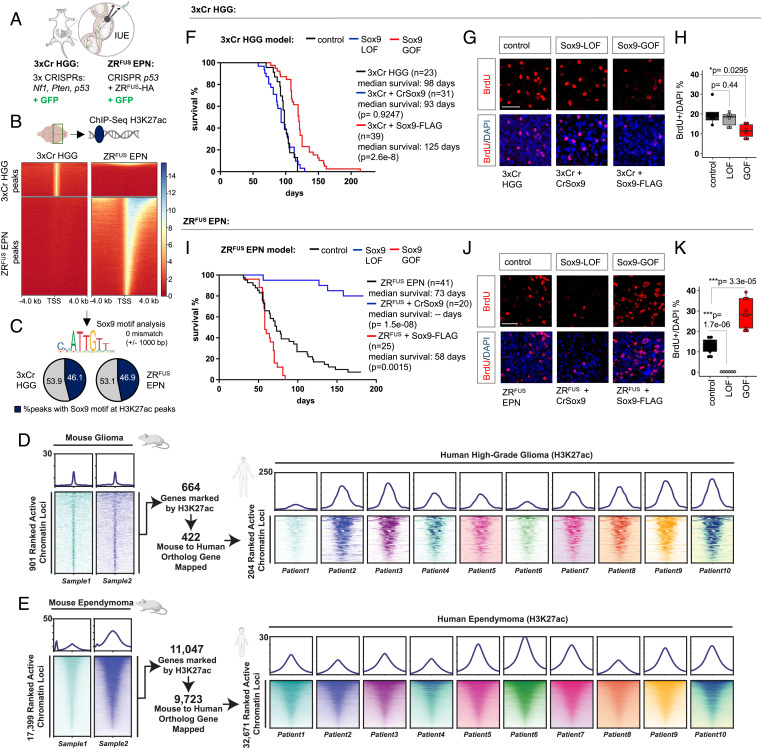
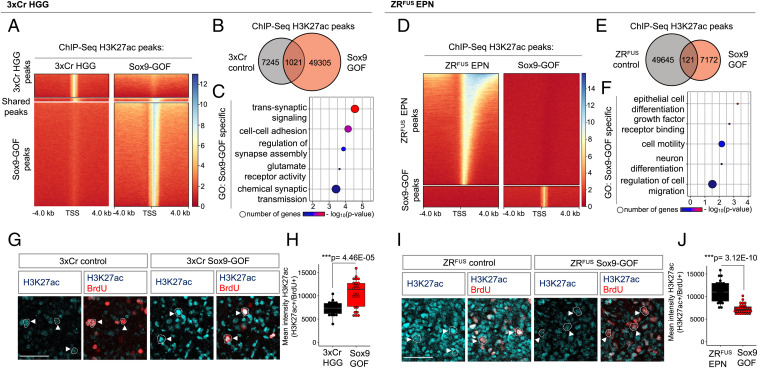
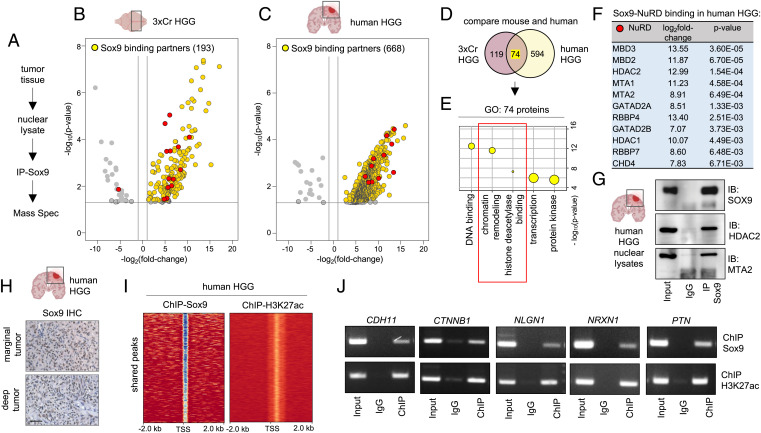
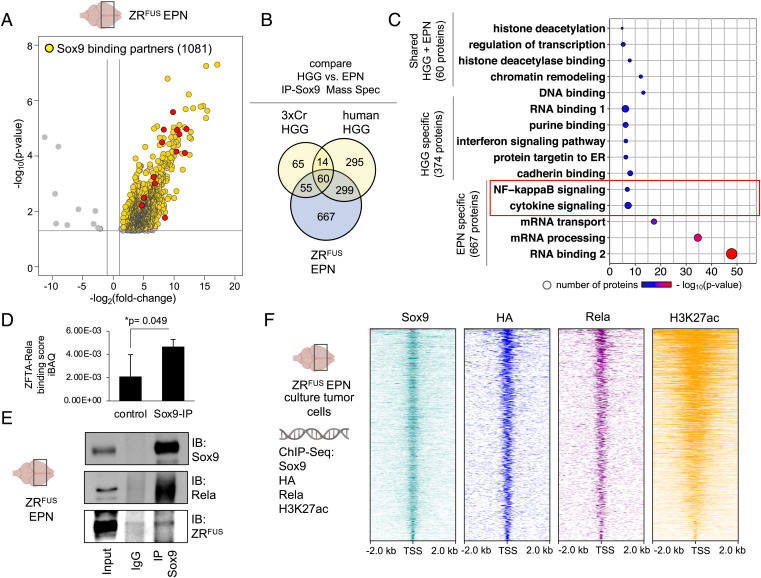
Epigenetic dysregulation is a universal feature of cancer that results in altered patterns of gene expression that drive malignancy. Brain tumors exhibit subtype-specific epigenetic alterations; however, the molecular mechanisms responsible for these diverse epigenetic states remain unclear. Here, we show that the developmental transcription factor Sox9 differentially regulates epigenomic states in high-grade glioma (HGG) and ependymoma (EPN). Using our autochthonous mouse models, we found that Sox9 suppresses HGG growth and expands associated H3K27ac states, while promoting ZFTA-RELA (ZRFUS) EPN growth and diminishing H3K27ac states. These contrasting roles for Sox9 correspond with protein interactions with histone deacetylating complexes in HGG and an association with the ZRFUS oncofusion in EPN. Mechanistic studies revealed extensive Sox9 and ZRFUS promoter co-occupancy, indicating functional synergy in promoting EPN tumorigenesis. Together, our studies demonstrate how epigenomic states are differentially regulated in distinct subtypes of brain tumors, while revealing divergent roles for Sox9 in HGG and EPN tumorigenesis.
Keywords: ependymoma; epigenetics; high-grade glioma; histone; transcription.
Conflict of interest statement
The authors declare no competing interest.
Figures

References
-
- Plass C., et al. , Mutations in regulators of the epigenome and their connections to global chromatin patterns in cancer. Nat. Rev. Genet. 14, 765–780 (2013). - PubMed
-
- Gallo M., et al. , MLL5 orchestrates a cancer self-renewal state by repressing the histone variant H3.3 and globally reorganizing chromatin. Cancer Cell 28, 715–729 (2015). - PubMed
-
- Schwartzentruber J., et al. , Driver mutations in histone H3.3 and chromatin remodelling genes in paediatric glioblastoma. Nature 482, 226–231 (2012). - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
