

PDGFRβ+ cells play a dual role as hematopoietic precursors and niche cells during mouse ontogeny
- PMID: 35858557
- PMCID: PMC9638014
- DOI: 10.1016/j.celrep.2022.111114
PDGFRβ+ cells play a dual role as hematopoietic precursors and niche cells during mouse ontogeny
Abstract
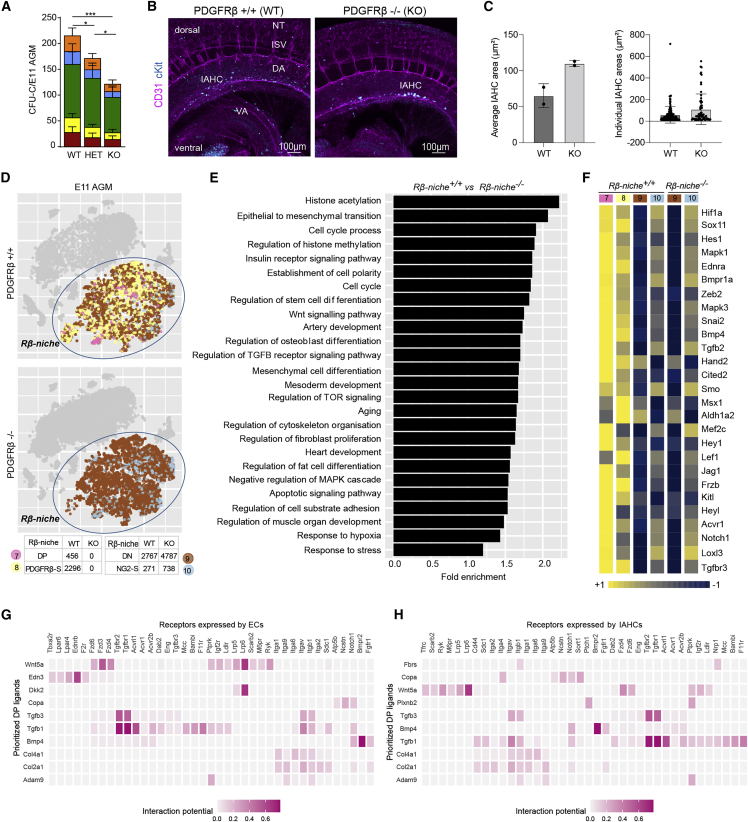
Hematopoietic stem cell (HSC) generation in the aorta-gonad-mesonephros region requires HSC specification signals from the surrounding microenvironment. In zebrafish, PDGF-B/PDGFRβ signaling controls hematopoietic stem/progenitor cell (HSPC) generation and is required in the HSC specification niche. Little is known about murine HSPC specification in vivo and whether PDGF-B/PDGFRβ is involved. Here, we show that PDGFRβ is expressed in distinct perivascular stromal cell layers surrounding the mid-gestation dorsal aorta, and its deletion impairs hematopoiesis. We demonstrate that PDGFRβ+ cells play a dual role in murine hematopoiesis. They act in the aortic niche to support HSPCs, and in addition, PDGFRβ+ embryonic precursors give rise to a subset of HSPCs that persist into adulthood. These findings provide crucial information for the controlled production of HSPCs in vitro.
Keywords: AGM single-cell RNA-sequencing; Bmp4; CP: Developmental biology; CP: Stem cell research; HSPC precursor; MSCs; PDGFRβ; VSMCs; hematopoietic niche; osteogenesis; pericytes.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures







Similar articles
-
A multistep computational approach reveals a neuro-mesenchymal cell population in the embryonic hematopoietic stem cell niche.Development. 2024 Apr 1;151(7):dev202614. doi: 10.1242/dev.202614. Epub 2024 Apr 4. Development. 2024. PMID: 38451068 Free PMC article.
-
Role of the microenvironment of the embryonic aorta-gonad-mesonephros region in hematopoiesis.Ann N Y Acad Sci. 2001 Jun;938:109-16. doi: 10.1111/j.1749-6632.2001.tb03579.x. Ann N Y Acad Sci. 2001. PMID: 11458497 Review.
-
HIF1α-induced PDGFRβ signaling promotes developmental HSC production via IL-6 activation.Exp Hematol. 2017 Feb;46:83-95.e6. doi: 10.1016/j.exphem.2016.10.002. Epub 2016 Oct 15. Exp Hematol. 2017. PMID: 27751871 Free PMC article.
-
Endothelium and NOTCH specify and amplify aorta-gonad-mesonephros-derived hematopoietic stem cells.J Clin Invest. 2015 May;125(5):2032-45. doi: 10.1172/JCI80137. Epub 2015 Apr 13. J Clin Invest. 2015. PMID: 25866967 Free PMC article.
-
Notch Signaling in HSC Emergence: When, Why and How.Cells. 2022 Jan 21;11(3):358. doi: 10.3390/cells11030358. Cells. 2022. PMID: 35159166 Free PMC article. Review.
Cited by
-
Sclerotome-derived PDGF signaling functions as a niche cue responsible for primitive erythropoiesis.Development. 2023 Nov 15;150(22):dev201807. doi: 10.1242/dev.201807. Epub 2023 Nov 16. Development. 2023. PMID: 37882745 Free PMC article.
-
The evolving hematopoietic niche during development.Front Mol Biosci. 2024 Oct 2;11:1488199. doi: 10.3389/fmolb.2024.1488199. eCollection 2024. Front Mol Biosci. 2024. PMID: 39417006 Free PMC article. Review.
-
A multistep computational approach reveals a neuro-mesenchymal cell population in the embryonic hematopoietic stem cell niche.Development. 2024 Apr 1;151(7):dev202614. doi: 10.1242/dev.202614. Epub 2024 Apr 4. Development. 2024. PMID: 38451068 Free PMC article.
-
The molecular and cellular hematopoietic stem cell specification niche.Exp Hematol. 2024 Aug;136:104280. doi: 10.1016/j.exphem.2024.104280. Epub 2024 Jul 14. Exp Hematol. 2024. PMID: 39009276 Free PMC article. Review.
-
Identification and characterization of enhancer elements controlling cell type-specific and signalling dependent chromatin programming during hematopoietic development.Stem Cell Investig. 2023 Jun 25;10:14. doi: 10.21037/sci-2023-011. eCollection 2023. Stem Cell Investig. 2023. PMID: 37404470 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
