

Alphaherpesvirus US3 protein-mediated inhibition of the m6A mRNA methyltransferase complex
- PMID: 35858564
- PMCID: PMC9347262
- DOI: 10.1016/j.celrep.2022.111107
Alphaherpesvirus US3 protein-mediated inhibition of the m6A mRNA methyltransferase complex
Abstract
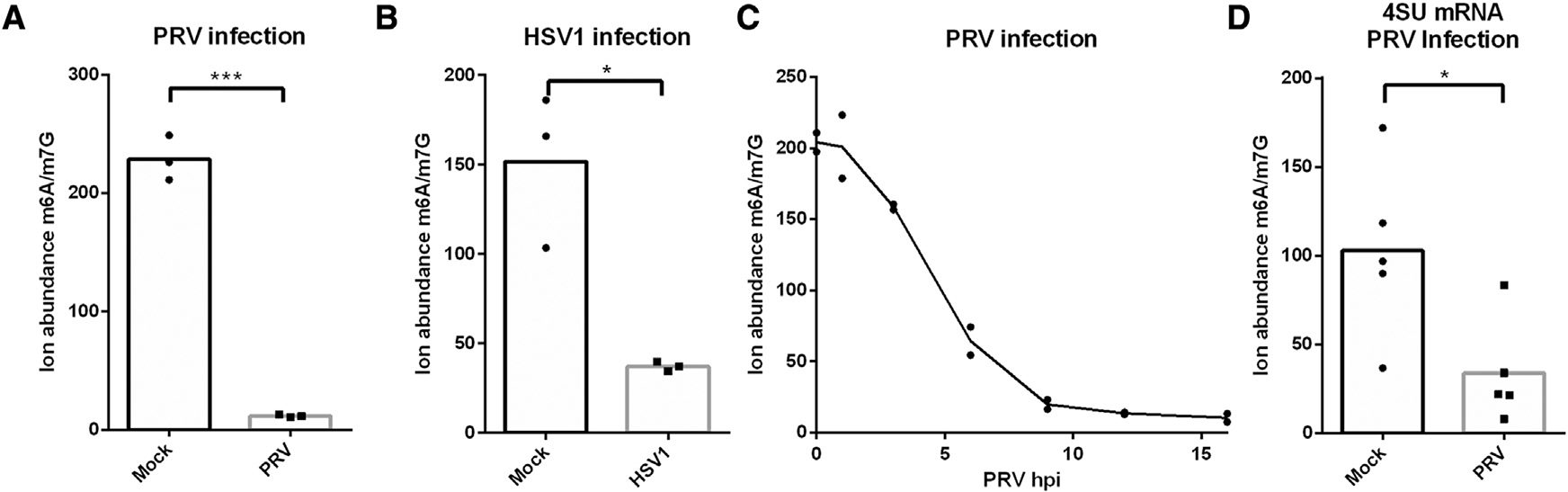
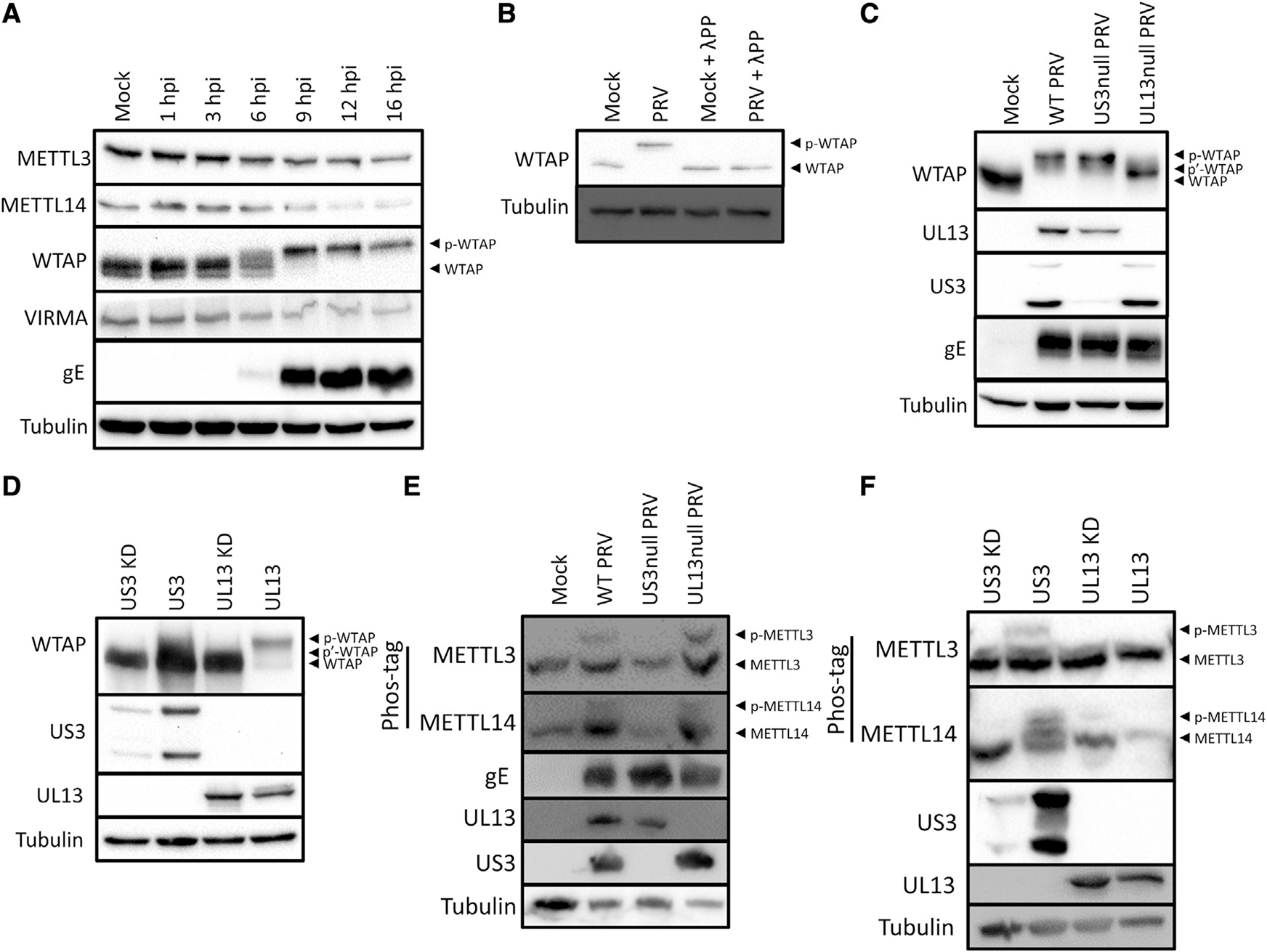
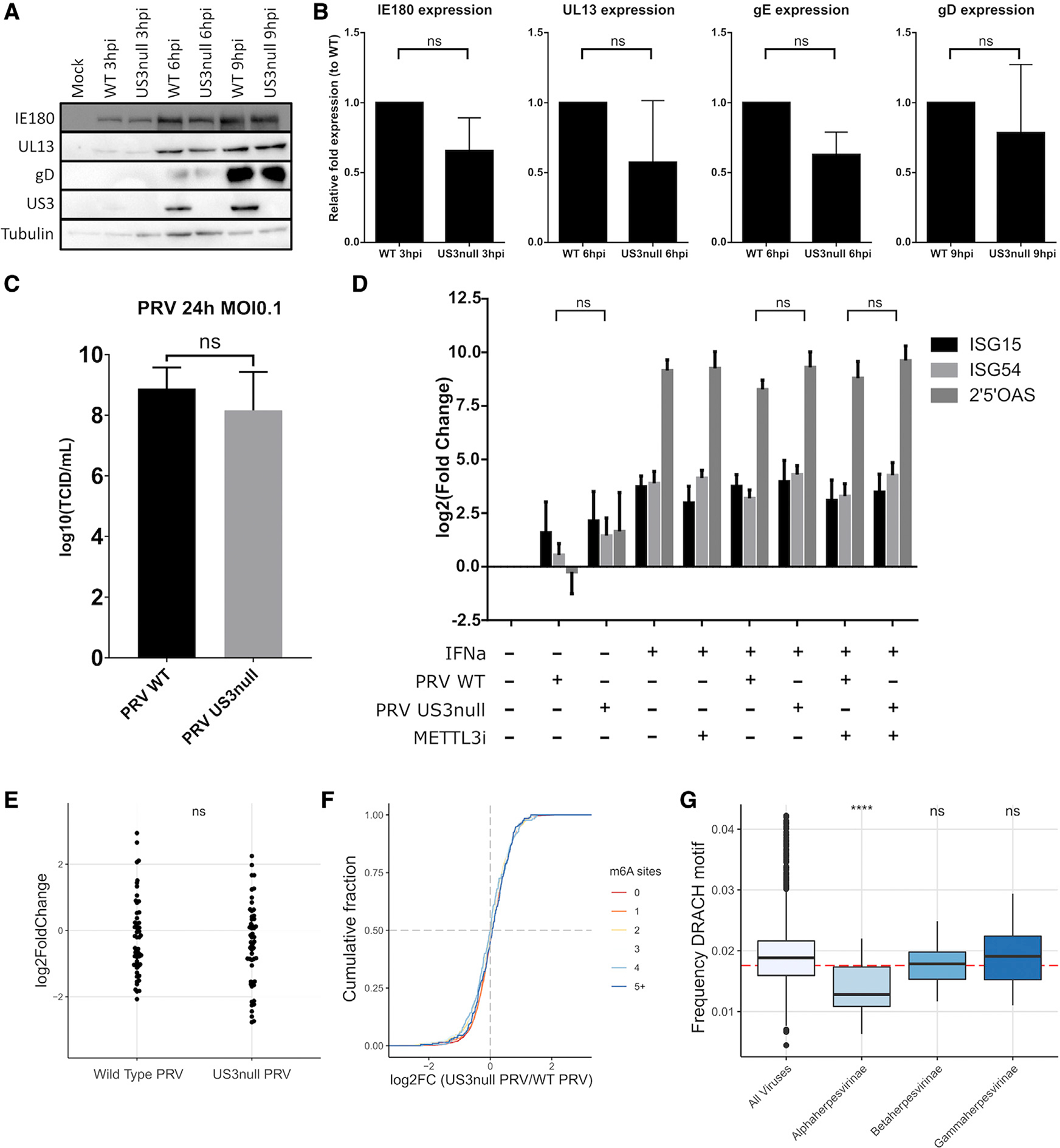
Chemical modifications of mRNA, the so-called epitranscriptome, represent an additional layer of post-transcriptional regulation of gene expression. The most common epitranscriptomic modification, N6-methyladenosine (m6A), is generated by a multi-subunit methyltransferase complex. We show that alphaherpesvirus kinases trigger phosphorylation of several components of the m6A methyltransferase complex, including METTL3, METTL14, and WTAP, which correlates with inhibition of the complex and a near complete loss of m6A levels in mRNA of virus-infected cells. Expression of the viral US3 protein is necessary and sufficient for phosphorylation and inhibition of the m6A methyltransferase complex. Although m6A methyltransferase complex inactivation is not essential for virus replication in cell culture, the consensus m6A methylation motif is under-represented in alphaherpesvirus genomes, suggesting evolutionary pressure against methylation of viral transcripts. Together, these findings reveal that phosphorylation can be associated with inactivation of the m6A methyltransferase complex, in this case mediated by the viral US3 protein.
Keywords: CP: Microbiology; CP: Molecular biology; PRV; US3; herpes; m6A; pseudorabies; writer complex.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.R.J. is an advisor to and owns equity in 858 Therapeutics and Lucerna Technologies.
Figures


References
-
- Bartkoski M, and Roizman B (1976). RNA synthesis in cells infected with herpes simple virus. XIII. Differences in the methylation patterns of viral RNA during the reproductive cycle. J. Virol. 20, 583–588. 10.1128/jvi.20.3.583-588.1976. http://www.ncbi.nlm.nih.gov/pubmed/186635. - DOI - PMC - PubMed
-
- Bokar JA, Rath-Shambaugh M, Ludwiczak R, Narayan P, and Rottman F (1994). Characterization and partial purification of mRNA N6-adenosine methyltransferase from HeLa cell nuclei. Internal mRNA methylation requires a multisubunit complex. J. Biol. Chem. 269, 17697–17704. 10.1016/s0021-9258(17)32497-3. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
