

Mucolytic bacteria license pathobionts to acquire host-derived nutrients during dietary nutrient restriction
- PMID: 35858565
- PMCID: PMC10903618
- DOI: 10.1016/j.celrep.2022.111093
Mucolytic bacteria license pathobionts to acquire host-derived nutrients during dietary nutrient restriction
Abstract
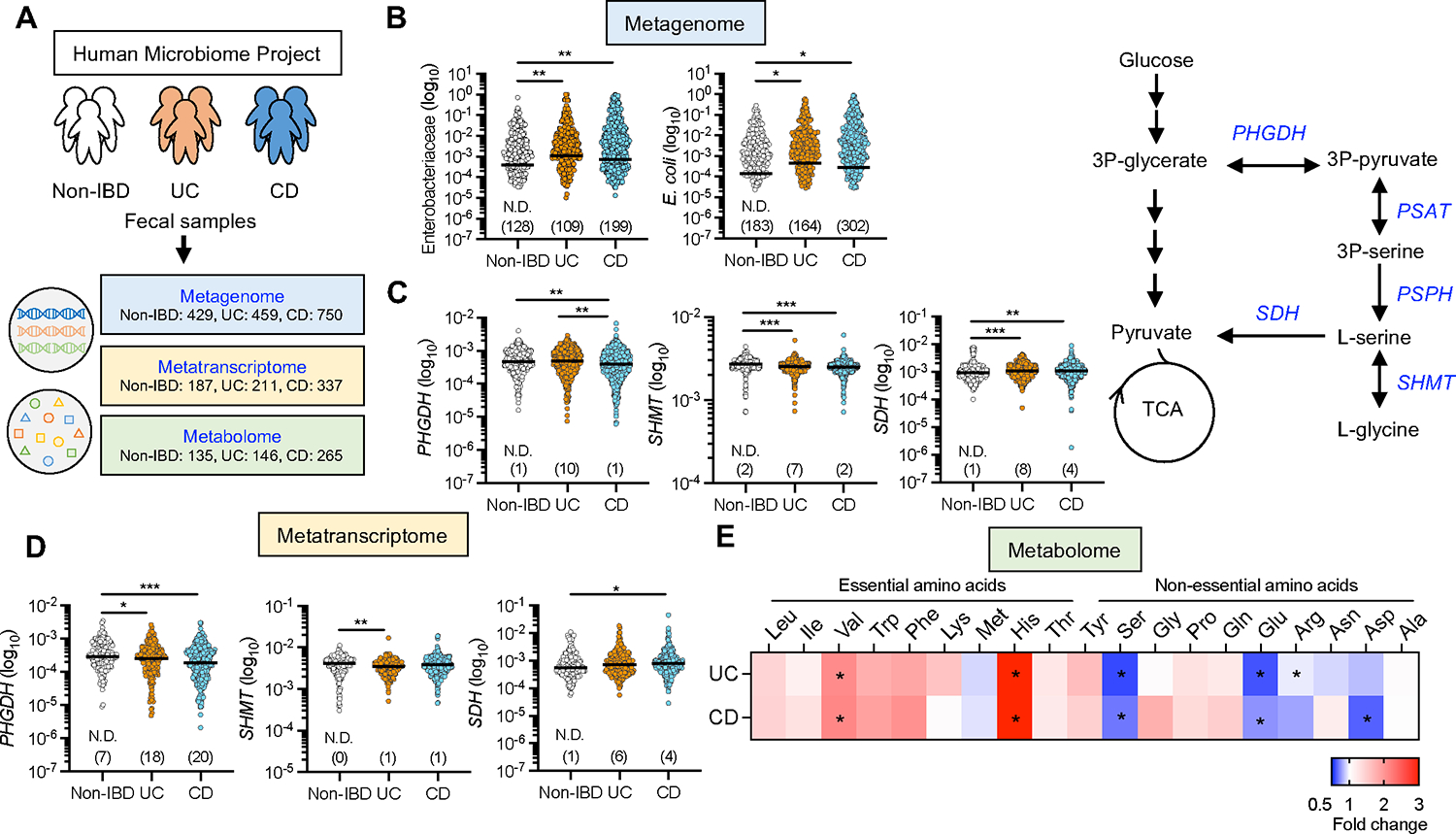
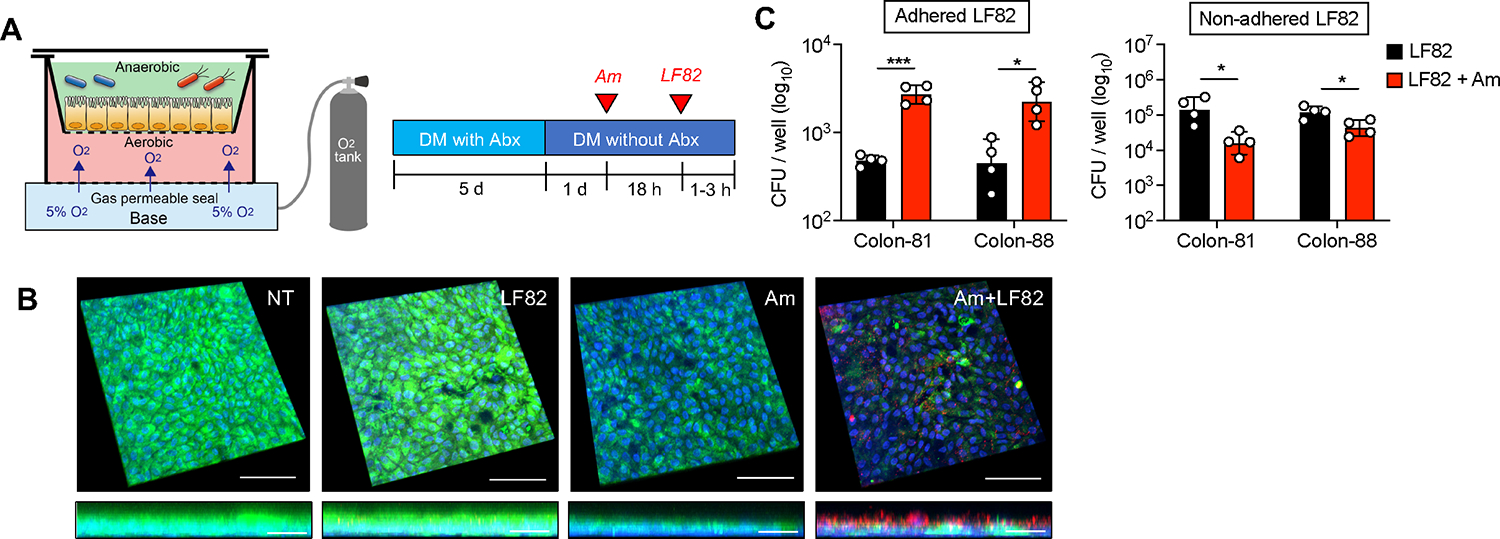
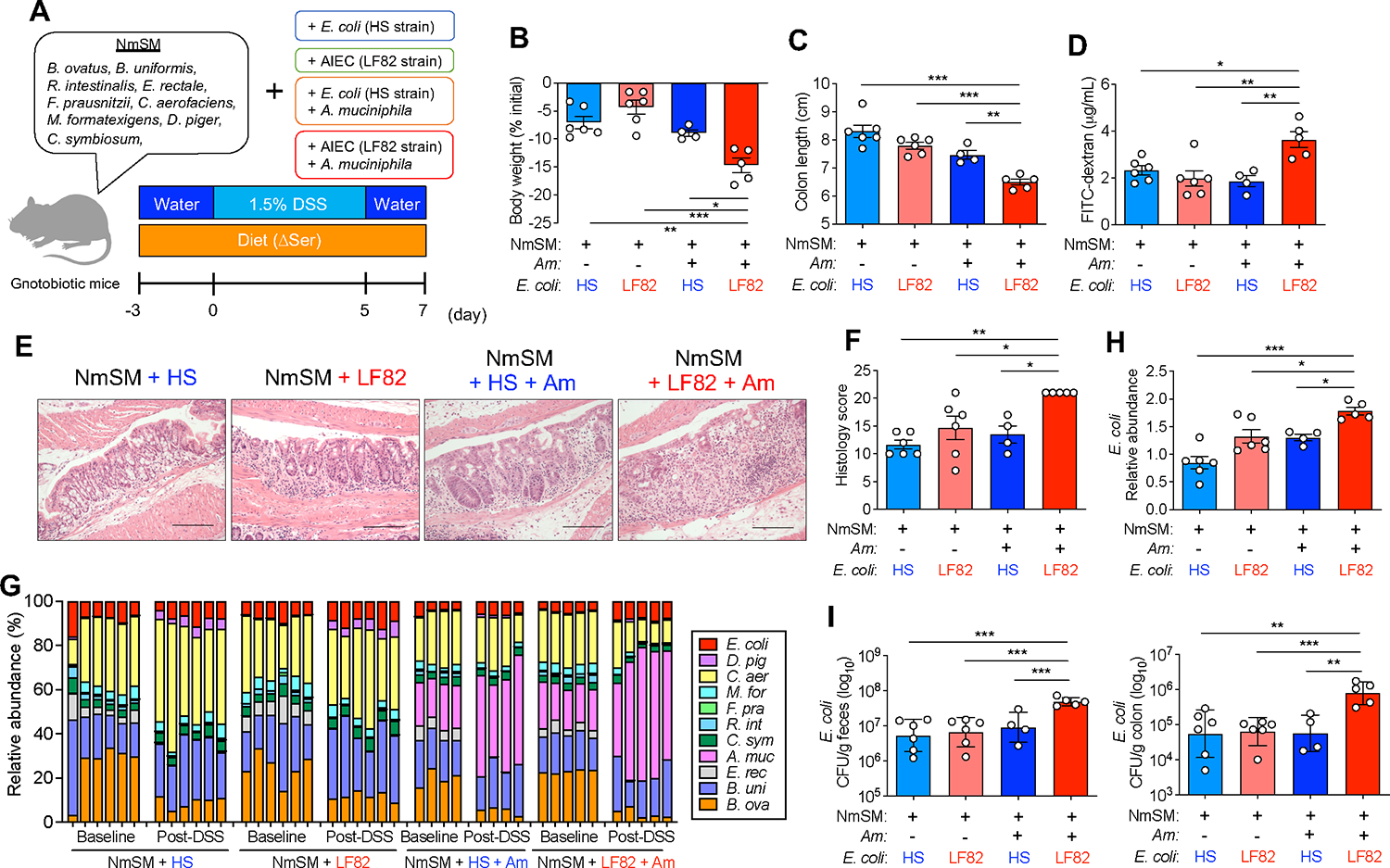
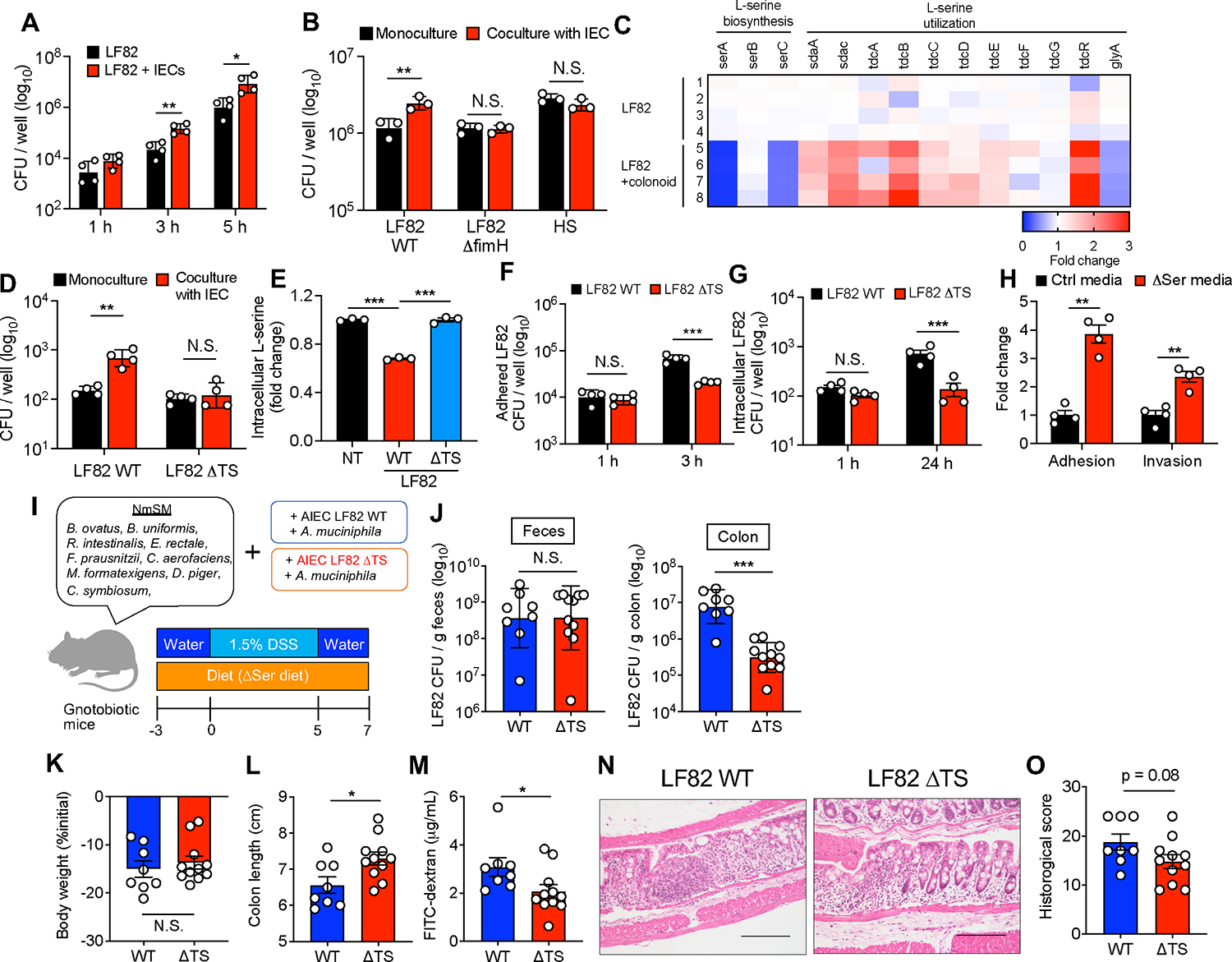
Pathobionts employ unique metabolic adaptation mechanisms to maximize their growth in disease conditions. Adherent-invasive Escherichia coli (AIEC), a pathobiont enriched in the gut mucosa of patients with inflammatory bowel disease (IBD), utilizes diet-derived L-serine to adapt to the inflamed gut. Therefore, the restriction of dietary L-serine starves AIEC and limits its fitness advantage. Here, we find that AIEC can overcome this nutrient limitation by switching the nutrient source from the diet to the host cells in the presence of mucolytic bacteria. During diet-derived L-serine restriction, the mucolytic symbiont Akkermansia muciniphila promotes the encroachment of AIEC to the epithelial niche by degrading the mucus layer. In the epithelial niche, AIEC acquires L-serine from the colonic epithelium and thus proliferates. Our work suggests that the indirect metabolic network between pathobionts and commensal symbionts enables pathobionts to overcome nutritional restriction and thrive in the gut.
Keywords: Akkermansia muciniphila; CP: Microbiology; L-serine; adherent-invasive Escherichia coli; inflammatory bowel disease; intestinal mucus barrier.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
