

Stress-induced perturbations in intracellular amino acids reprogram mRNA translation in osmoadaptation independently of the ISR
- PMID: 35858571
- PMCID: PMC9491157
- DOI: 10.1016/j.celrep.2022.111092
Stress-induced perturbations in intracellular amino acids reprogram mRNA translation in osmoadaptation independently of the ISR
Abstract
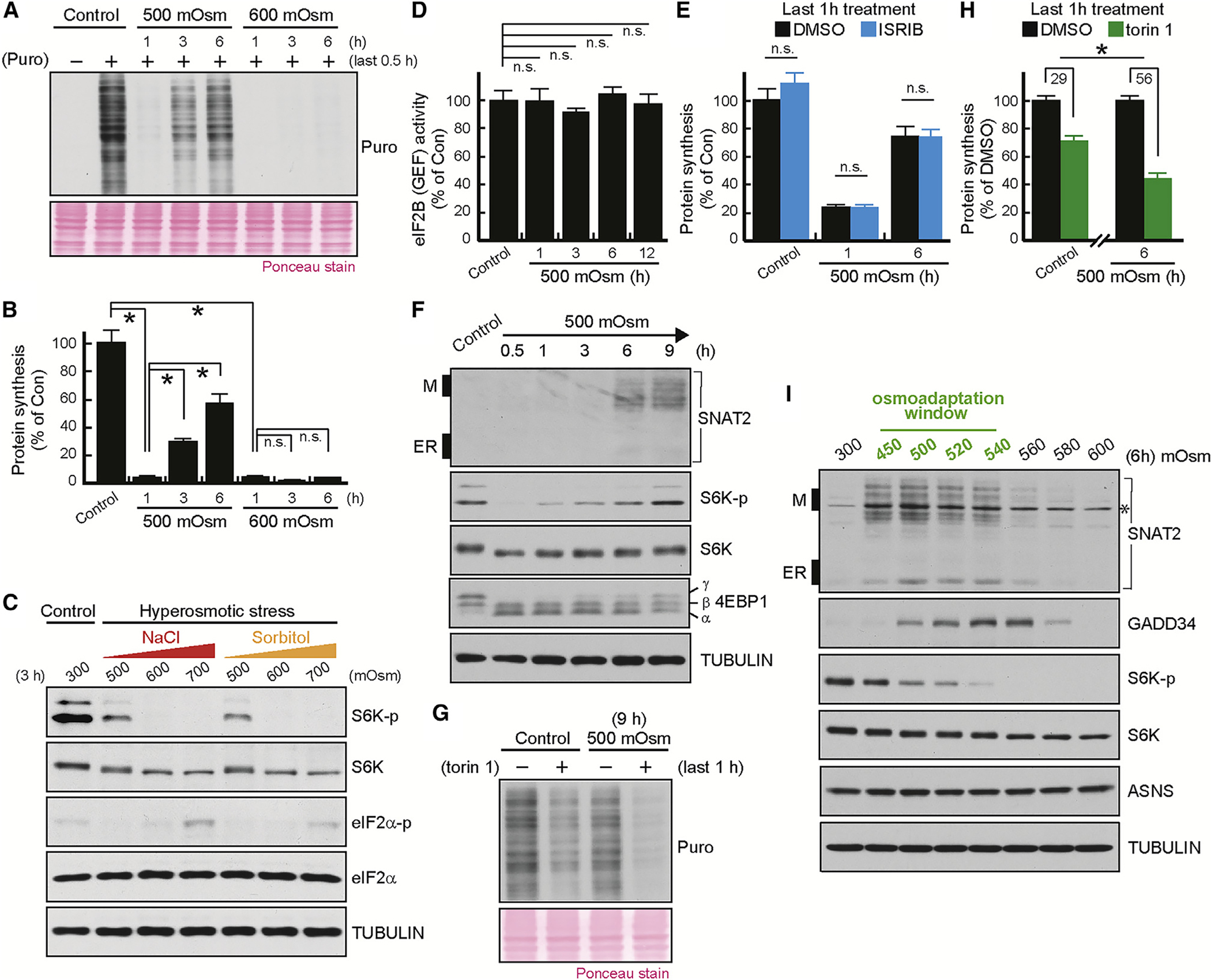
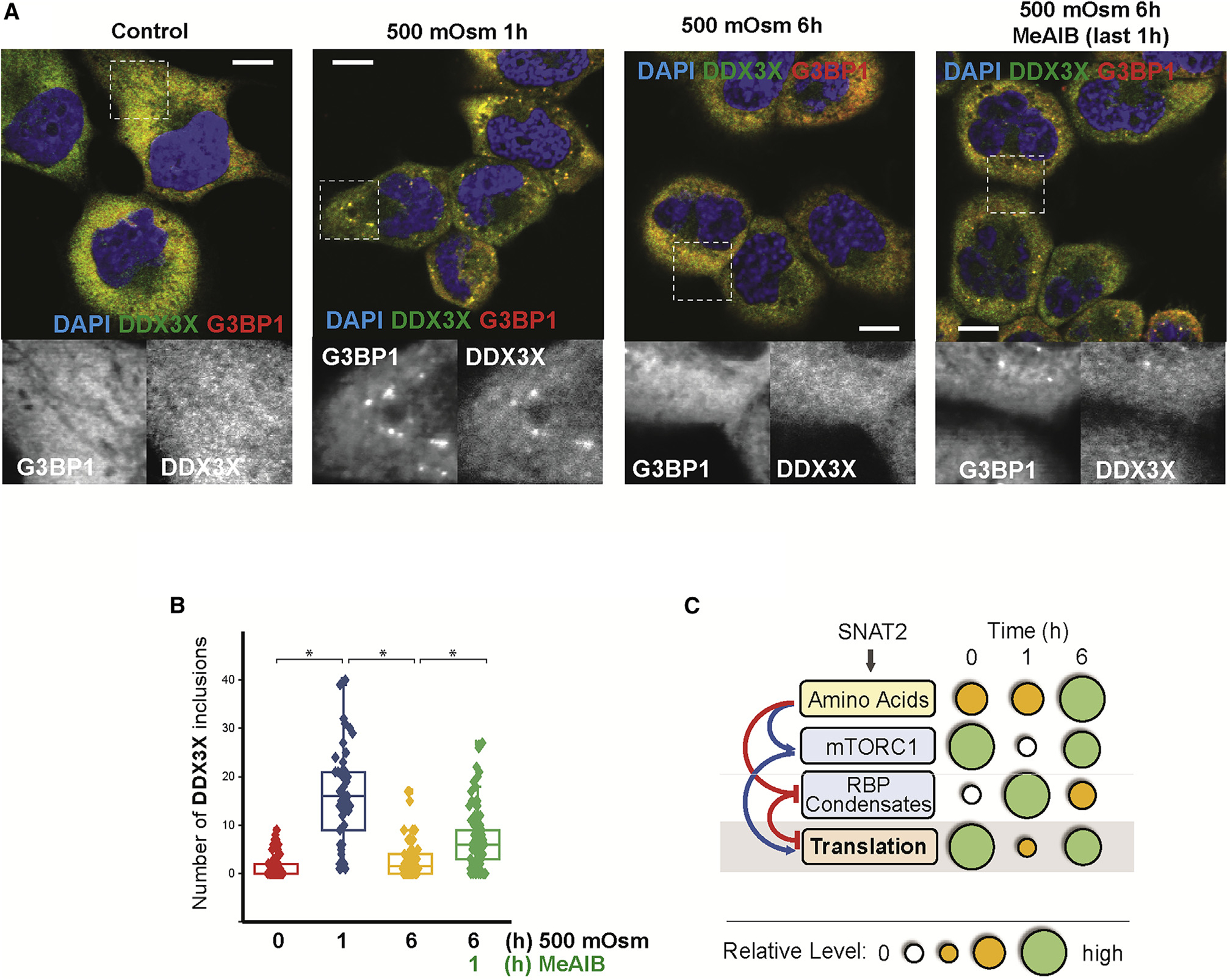
The integrated stress response (ISR) plays a pivotal role in adaptation of translation machinery to cellular stress. Here, we demonstrate an ISR-independent osmoadaptation mechanism involving reprogramming of translation via coordinated but independent actions of mTOR and plasma membrane amino acid transporter SNAT2. This biphasic response entails reduced global protein synthesis and mTOR signaling followed by translation of SNAT2. Induction of SNAT2 leads to accumulation of amino acids and reactivation of mTOR and global protein synthesis, paralleled by partial reversal of the early-phase, stress-induced translatome. We propose SNAT2 functions as a molecular switch between inhibition of protein synthesis and establishment of an osmoadaptive translation program involving the formation of cytoplasmic condensates of SNAT2-regulated RNA-binding proteins DDX3X and FUS. In summary, we define key roles of SNAT2 in osmotolerance.
Keywords: CP: Molecular biology; amino acids; cytoplasmic condensates; hypertonic stress; mTOR signaling; osmolytes; osmotolerance; translation.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
- T34 GM137792/GM/NIGMS NIH HHS/United States
- R37 DK060596/DK/NIDDK NIH HHS/United States
- P30 DK097948/DK/NIDDK NIH HHS/United States
- R25 GM049010/GM/NIGMS NIH HHS/United States
- R01 DK060596/DK/NIDDK NIH HHS/United States
- R01 GM128981/GM/NIGMS NIH HHS/United States
- R01 NS117176/NS/NINDS NIH HHS/United States
- F30 CA247347/CA/NCI NIH HHS/United States
- K01 NS116119/NS/NINDS NIH HHS/United States
- T32 GM007250/GM/NIGMS NIH HHS/United States
- R35 GM118088/GM/NIGMS NIH HHS/United States
- BBS/E/B/000C0413/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- R01 NS105789/NS/NINDS NIH HHS/United States
- R01 DK053307/DK/NIDDK NIH HHS/United States
- R01 NS121374/NS/NINDS NIH HHS/United States
- R01 DK113196/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
