

Ablation of Siglec-E augments brain inflammation and ischemic injury
- PMID: 35858866
- PMCID: PMC9301848
- DOI: 10.1186/s12974-022-02556-1
Ablation of Siglec-E augments brain inflammation and ischemic injury
Abstract
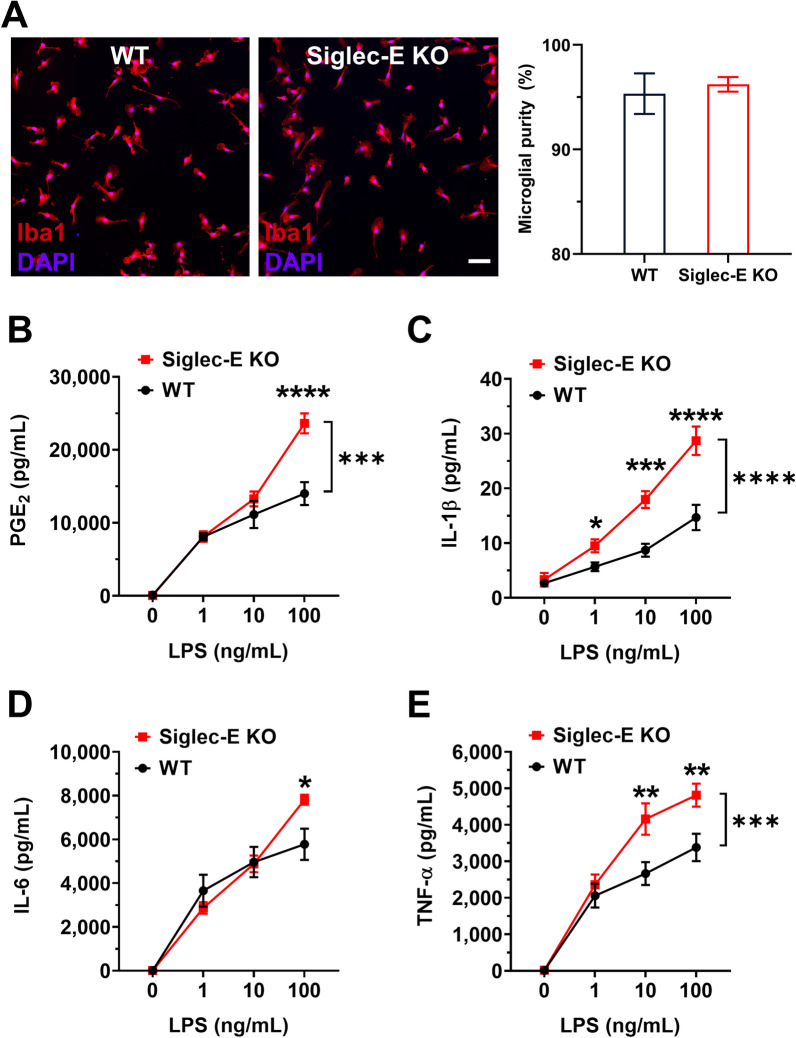
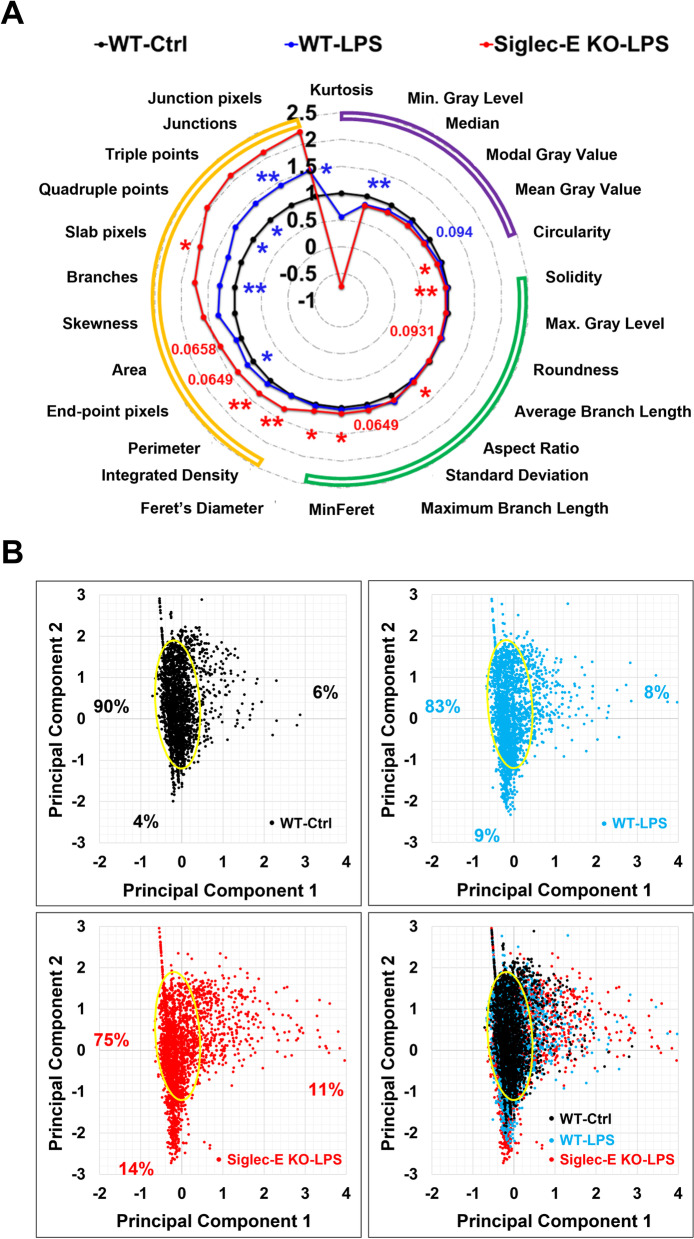
Sialic acid immunoglobulin-like lectin E (Siglec-E) is a subtype of pattern recognition receptors found on the surface of myeloid cells and functions as a key immunosuppressive checkpoint molecule. The engagement between Siglec-E and the ligand α2,8-linked disialyl glycans activates the immunoreceptor tyrosine-based inhibitory motif (ITIM) in its intracellular domain, mitigating the potential risk of autoimmunity amid innate immune attacks on parasites, bacteria, and carcinoma. Recent studies suggest that Siglec-E is also expressed in the CNS, particularly microglia, the brain-resident immune cells. However, the functions of Siglec-E in brain inflammation and injuries under many neurological conditions largely remain elusive. In this study, we first revealed an anti-inflammatory role for Siglec-E in lipopolysaccharide (LPS)-triggered microglial activation. We then found that Siglec-E was induced within the brain by systemic treatment with LPS in mice in a dose-dependent manner, while its ablation exacerbated hippocampal reactive microgliosis in LPS-treated animals. The genetic deficiency of Siglec-E also aggravated oxygen-glucose deprivation (OGD)-induced neuronal death in mouse primary cortical cultures containing both neurons and glial cells. Moreover, Siglec-E expression in ipsilateral brain tissues was substantially induced following middle cerebral artery occlusion (MCAO). Lastly, the neurological deficits and brain infarcts were augmented in Siglec-E knockout mice after moderate MCAO when compared to wild-type animals. Collectively, our findings suggest that the endogenous inducible Siglec-E plays crucial anti-inflammatory and neuroprotective roles following ischemic stroke, and thus might underlie an intrinsic mechanism of resolution of inflammation and self-repair in the brain.
Keywords: Cerebral ischemia; DAMPs; Infarct; Innate immunity; LPS; Microgliosis; Neuroprotection; OGD; PAMPs; Sialic acid.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
The authors declare that they have no competing interests.
Figures


Similar articles
-
Sensing the neuronal glycocalyx by glial sialic acid binding immunoglobulin-like lectins.Neuroscience. 2014 Sep 5;275:113-24. doi: 10.1016/j.neuroscience.2014.05.061. Epub 2014 Jun 9. Neuroscience. 2014. PMID: 24924144 Review.
-
Immunosuppressive Siglec-E ligands on mouse aorta are up-regulated by LPS via NF-κB pathway.Biomed Pharmacother. 2020 Feb;122:109760. doi: 10.1016/j.biopha.2019.109760. Epub 2019 Dec 30. Biomed Pharmacother. 2020. PMID: 31918287
-
Alleviation of neurotoxicity by microglial human Siglec-11.J Neurosci. 2010 Mar 3;30(9):3482-8. doi: 10.1523/JNEUROSCI.3940-09.2010. J Neurosci. 2010. PMID: 20203208 Free PMC article.
-
Siglec functions of microglia.Glycobiology. 2014 Sep;24(9):794-9. doi: 10.1093/glycob/cwu044. Epub 2014 May 15. Glycobiology. 2014. PMID: 24833613 Review.
-
Phosphoproteomics identifies microglial Siglec-F inflammatory response during neurodegeneration.Mol Syst Biol. 2020 Dec;16(12):e9819. doi: 10.15252/msb.20209819. Mol Syst Biol. 2020. PMID: 33289969 Free PMC article.
Cited by
-
Siglec-9 is an inhibitory receptor on human mast cells in vitro.J Allergy Clin Immunol. 2023 Sep;152(3):711-724.e14. doi: 10.1016/j.jaci.2023.04.007. Epub 2023 Apr 24. J Allergy Clin Immunol. 2023. PMID: 37100120 Free PMC article.
-
Regulators of phagocytosis as pharmacologic targets for stroke treatment.Front Pharmacol. 2023 Aug 2;14:1122527. doi: 10.3389/fphar.2023.1122527. eCollection 2023. Front Pharmacol. 2023. PMID: 37601043 Free PMC article. Review.
-
Fatty Acid-Binding Protein 4 is Essential for the Inflammatory and Metabolic Response of Microglia to Lipopolysaccharide.J Neuroimmune Pharmacol. 2023 Sep;18(3):448-461. doi: 10.1007/s11481-023-10079-6. Epub 2023 Aug 9. J Neuroimmune Pharmacol. 2023. PMID: 37555918 Free PMC article.
-
Therapeutic potential to target sialylation and SIGLECs in neurodegenerative and psychiatric diseases.Front Neurol. 2024 Mar 11;15:1330874. doi: 10.3389/fneur.2024.1330874. eCollection 2024. Front Neurol. 2024. PMID: 38529039 Free PMC article. Review.
-
Siglec-E Ligand Downregulation on Hippocampus Neurons Induced Inflammation in Sevoflurane-Associated Perioperative Neurocognitive Disorders in Aged Mice.Inflammation. 2024 Feb;47(1):30-44. doi: 10.1007/s10753-023-01888-1. Epub 2023 Aug 21. Inflammation. 2024. PMID: 37603227
References
-
- Varki A, Schnaar RL, Crocker PR, et al. I-type lectins. In: Varki A, Cummings RD, Esko JD, Stanley P, Hart GW, Aebi M, Darvill AG, Kinoshita T, Packer NH, Prestegard JH, et al., editors. Essentials of glycobiology. 3. La Jolla: Cold Spring Harbor Laboratory Press; 2015. pp. 453–467. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
