

Structure and specialization of mycorrhizal networks in phylogenetically diverse tropical communities
- PMID: 35859141
- PMCID: PMC9297633
- DOI: 10.1186/s40793-022-00434-0
Structure and specialization of mycorrhizal networks in phylogenetically diverse tropical communities
Abstract
Background: The root mycobiome plays a fundamental role in plant nutrition and protection against biotic and abiotic stresses. In temperate forests or meadows dominated by angiosperms, the numerous fungi involved in root symbioses are often shared between neighboring plants, thus forming complex plant-fungus interaction networks of weak specialization. Whether this weak specialization also holds in rich tropical communities with more phylogenetically diverse sets of plant lineages remains unknown. We collected roots of 30 plant species in semi-natural tropical communities including angiosperms, ferns, and lycophytes, in three different habitat types on La Réunion island: a recent lava flow, a wet thicket, and an ericoid shrubland. We identified root-inhabiting fungi by sequencing both the 18S rRNA and the ITS2 variable regions. We assessed the diversity of mycorrhizal fungal taxa according to plant species and lineages, as well as the structure and specialization of the resulting plant-fungus networks.
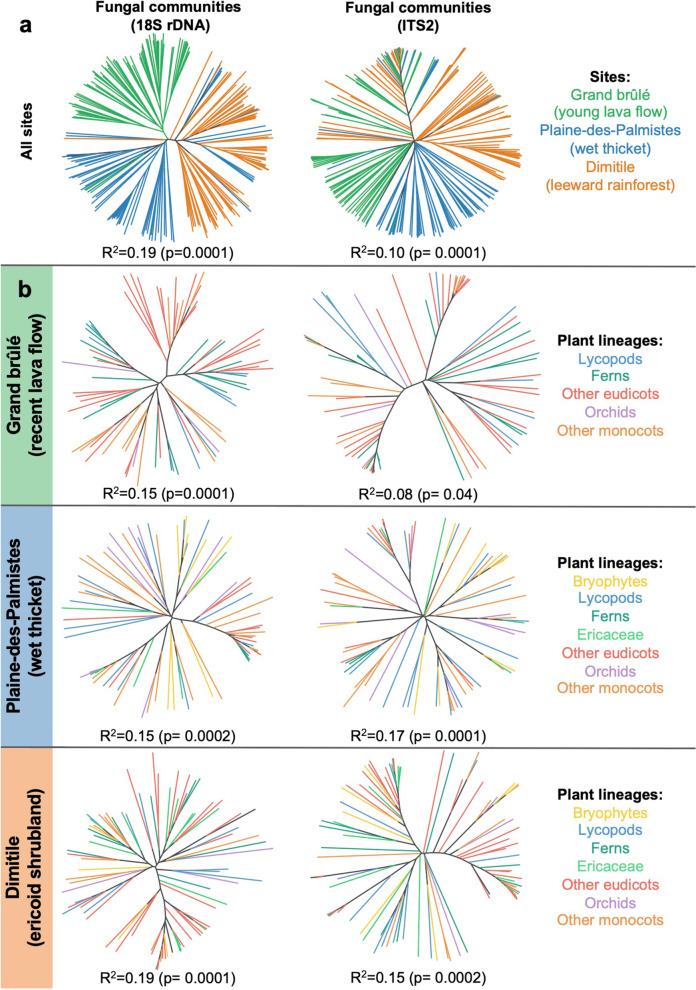
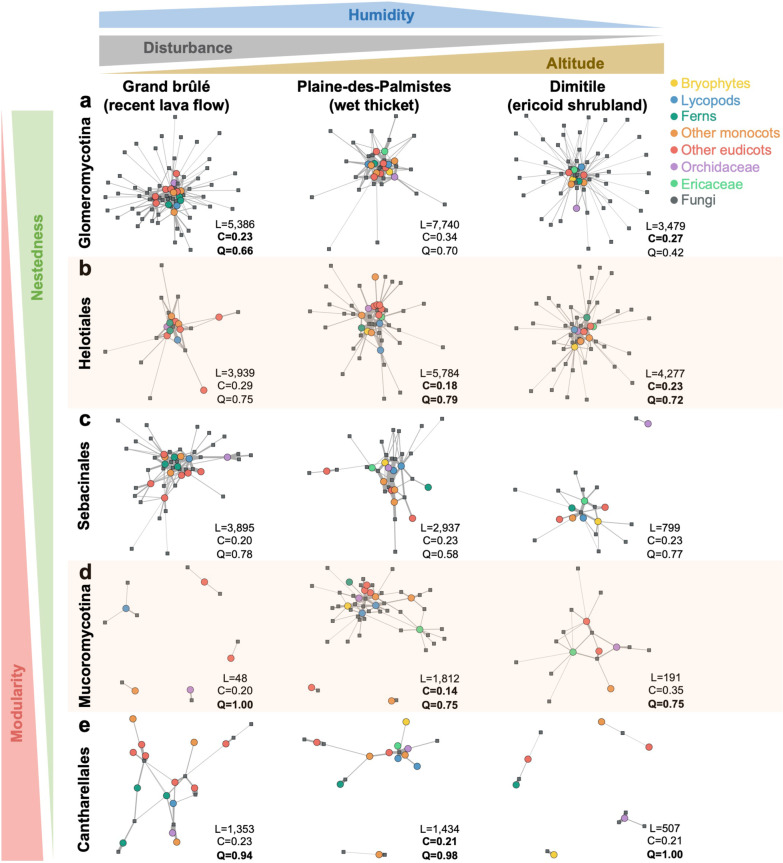
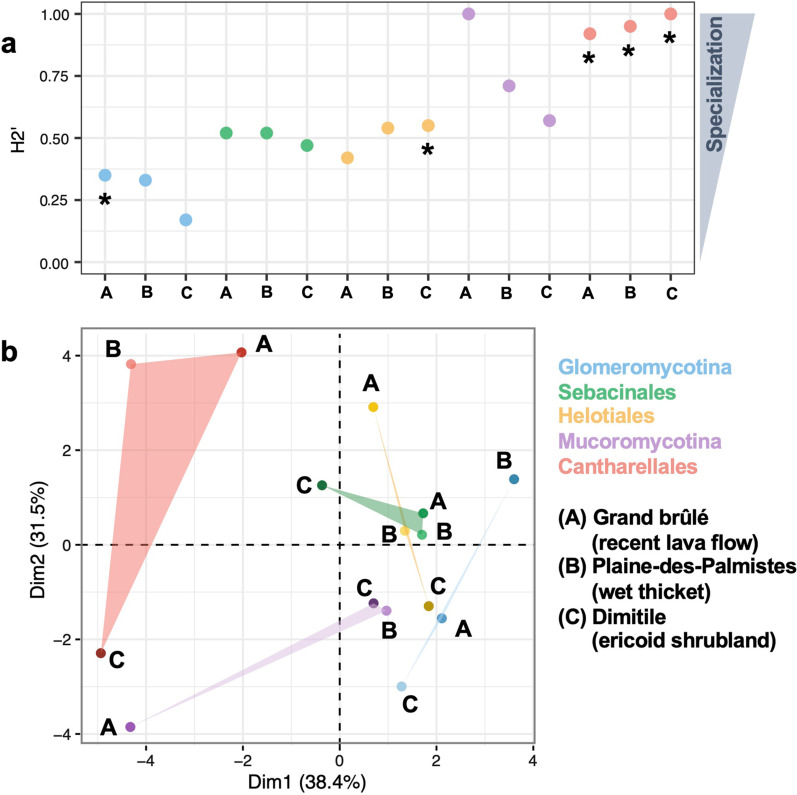
Results: The 18S and ITS2 datasets are highly complementary at revealing the root mycobiota. According to 18S, Glomeromycotina colonize all plant groups in all habitats forming the least specialized interactions, resulting in nested network structures, while Mucoromycotina (Endogonales) are more abundant in the wetland and show higher specialization and modularity compared to the former. According to ITS2, mycorrhizal fungi of Ericaceae and Orchidaceae, namely Helotiales, Sebacinales, and Cantharellales, also colonize the roots of most plant lineages, confirming that they are frequent endophytes. While Helotiales and Sebacinales present intermediate levels of specialization, Cantharellales are more specialized and more sporadic in their interactions with plants, resulting in highly modular networks.
Conclusions: This study of the root mycobiome in tropical environments reinforces the idea that mycorrhizal fungal taxa are locally shared between co-occurring plants, including phylogenetically distant plants (e.g. lycophytes and angiosperms), where they may form functional mycorrhizae or establish endophytic colonization. Yet, we demonstrate that, irrespectively of the environmental variations, the level of specialization significantly varies according to the fungal lineages, probably reflecting the different evolutionary origins of these plant-fungus symbioses. Frequent fungal sharing between plants questions the roles of the different fungi in community functioning and highlights the importance of considering networks of interactions rather than isolated hosts.
Keywords: Common mycorrhizal networks; Early-diverging plants; Endomycorrhiza; Fungal metabarcoding; Mucoromycotina fine root endophytes; Root endophytism.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interest.
Figures


Similar articles
-
Needle in a Haystack: Culturing Plant-Beneficial Helotiales Lineages From Plant Roots.Environ Microbiol. 2025 Apr;27(4):e70082. doi: 10.1111/1462-2920.70082. Environ Microbiol. 2025. PMID: 40228525 Free PMC article.
-
Sharing of diverse mycorrhizal and root-endophytic fungi among plant species in an oak-dominated cool-temperate forest.PLoS One. 2013 Oct 21;8(10):e78248. doi: 10.1371/journal.pone.0078248. eCollection 2013. PLoS One. 2013. PMID: 24250752 Free PMC article.
-
Diversity and spatial structure of belowground plant-fungal symbiosis in a mixed subtropical forest of ectomycorrhizal and arbuscular mycorrhizal plants.PLoS One. 2014 Jan 28;9(1):e86566. doi: 10.1371/journal.pone.0086566. eCollection 2014. PLoS One. 2014. PMID: 24489745 Free PMC article.
-
Unity in diversity: structural and functional insights into the ancient partnerships between plants and fungi.New Phytol. 2018 Dec;220(4):996-1011. doi: 10.1111/nph.15158. Epub 2018 Apr 26. New Phytol. 2018. PMID: 29696662 Review.
-
The Waiting Room Hypothesis revisited by orchids: were orchid mycorrhizal fungi recruited among root endophytes?Ann Bot. 2022 Feb 11;129(3):259-270. doi: 10.1093/aob/mcab134. Ann Bot. 2022. PMID: 34718377 Free PMC article. Review.
Cited by
-
Fungal microbiomes associated with Lycopodiaceae during ecological succession.Environ Microbiol Rep. 2023 Apr;15(2):109-118. doi: 10.1111/1758-2229.13130. Epub 2022 Oct 10. Environ Microbiol Rep. 2023. PMID: 36216403 Free PMC article.
-
Assessing the structure and diversity of fungal community in plant soil under different climatic and vegetation conditions.Front Microbiol. 2023 Nov 29;14:1288066. doi: 10.3389/fmicb.2023.1288066. eCollection 2023. Front Microbiol. 2023. PMID: 38094633 Free PMC article.
-
A metabarcoding protocol targeting two DNA regions to analyze root-associated fungal communities in ferns and lycophytes.Appl Plant Sci. 2023 Jun 12;11(3):e11523. doi: 10.1002/aps3.11523. eCollection 2023 May-Jun. Appl Plant Sci. 2023. PMID: 37342167 Free PMC article.
-
Comparative analysis of mycorrhizal communities associated with Struthiopteris spicant (L.) Weiss across Europe and North America.Front Plant Sci. 2024 Jun 4;15:1402946. doi: 10.3389/fpls.2024.1402946. eCollection 2024. Front Plant Sci. 2024. PMID: 38899157 Free PMC article.
-
Seed or soil: Tracing back the plant mycobiota primary sources.Environ Microbiol Rep. 2024 Jun;16(3):e13301. doi: 10.1111/1758-2229.13301. Environ Microbiol Rep. 2024. PMID: 38924368 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources