

Sestrin mediates detection of and adaptation to low-leucine diets in Drosophila
- PMID: 35859173
- PMCID: PMC10112710
- DOI: 10.1038/s41586-022-04960-2
Sestrin mediates detection of and adaptation to low-leucine diets in Drosophila
Erratum in
-
Author Correction: Sestrin mediates detection of and adaptation to low-leucine diets in Drosophila.Nature. 2022 Sep;609(7929):E11. doi: 10.1038/s41586-022-05286-9. Nature. 2022. PMID: 36100671 No abstract available.
Abstract
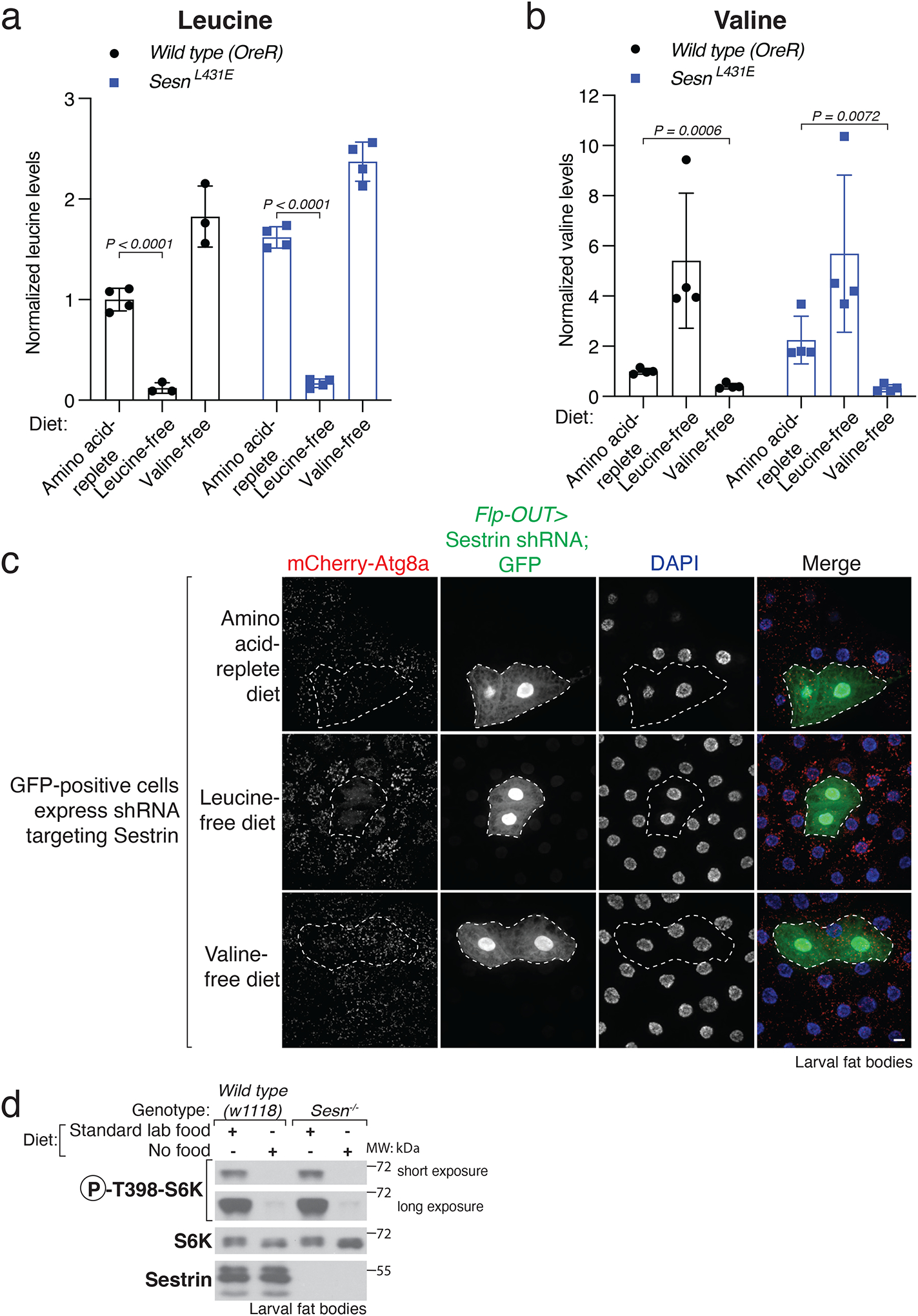
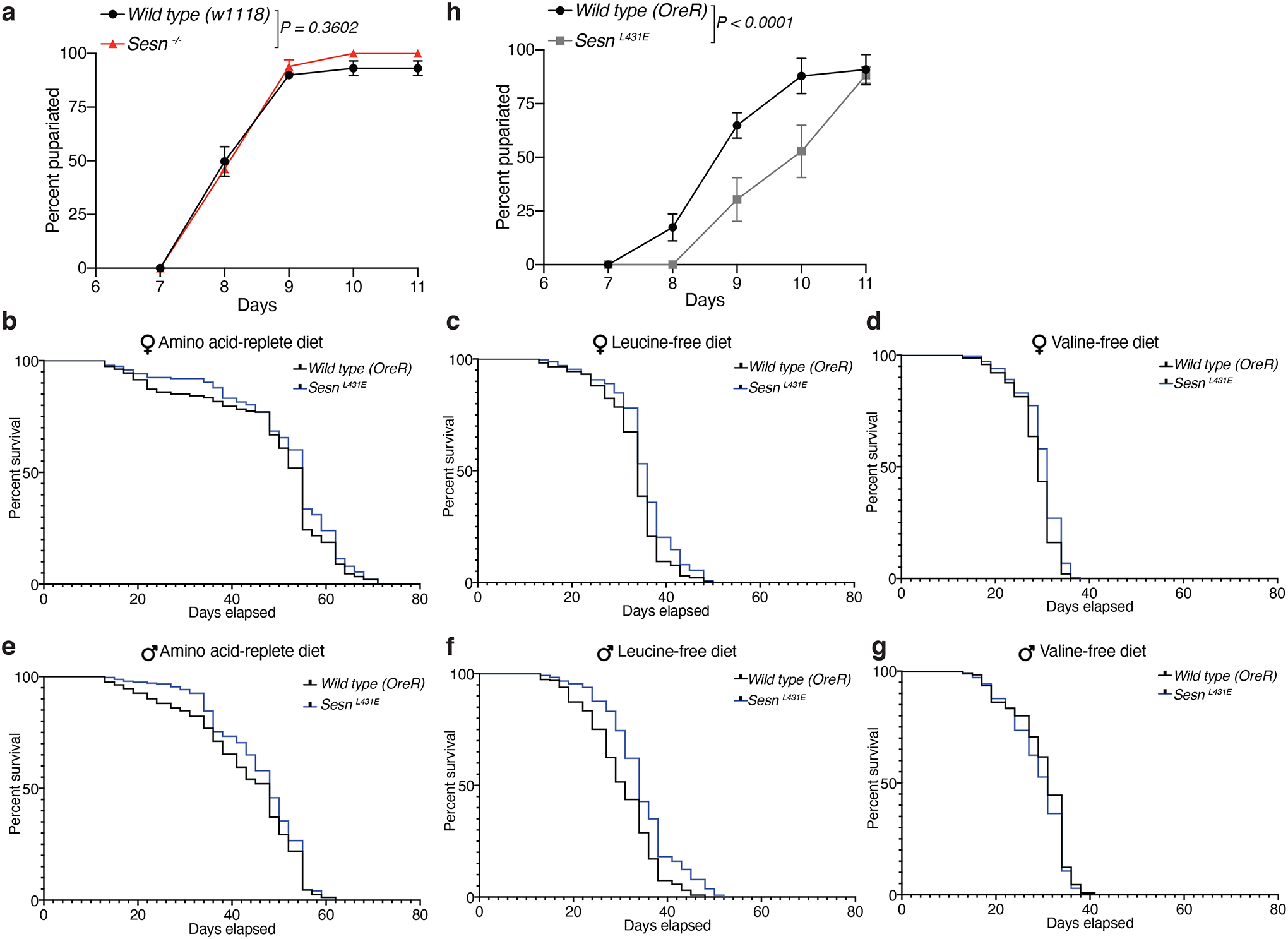
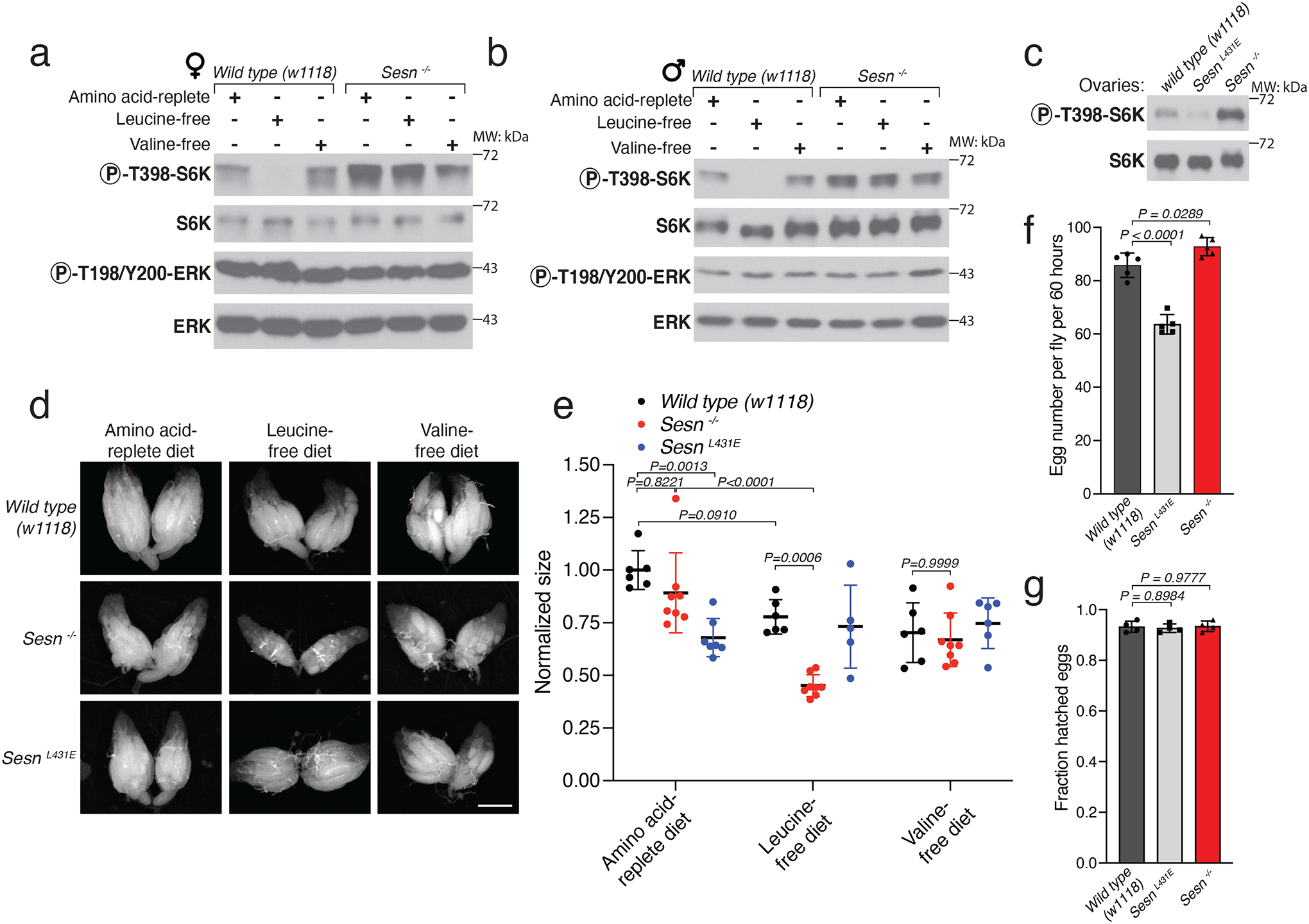
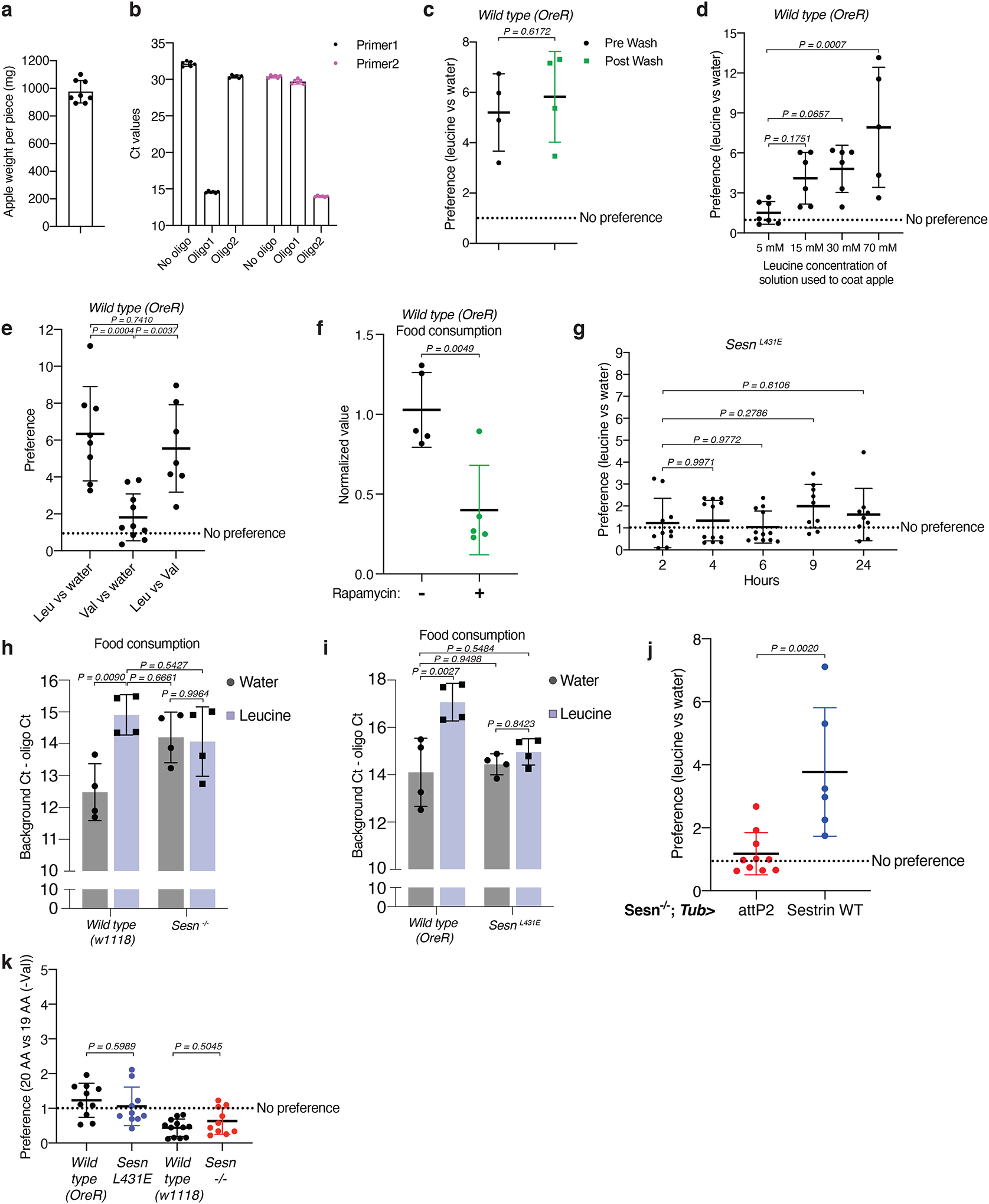
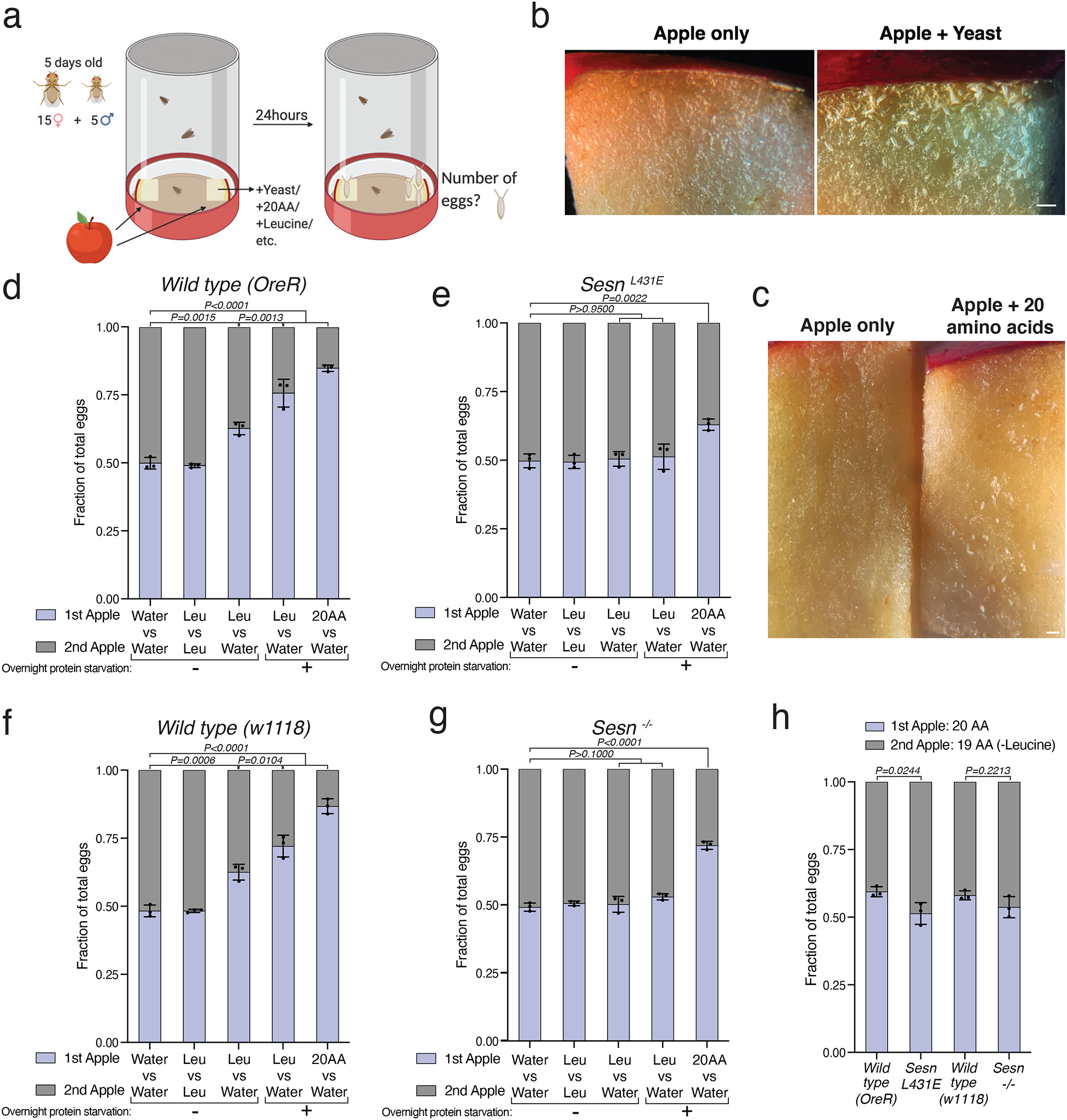
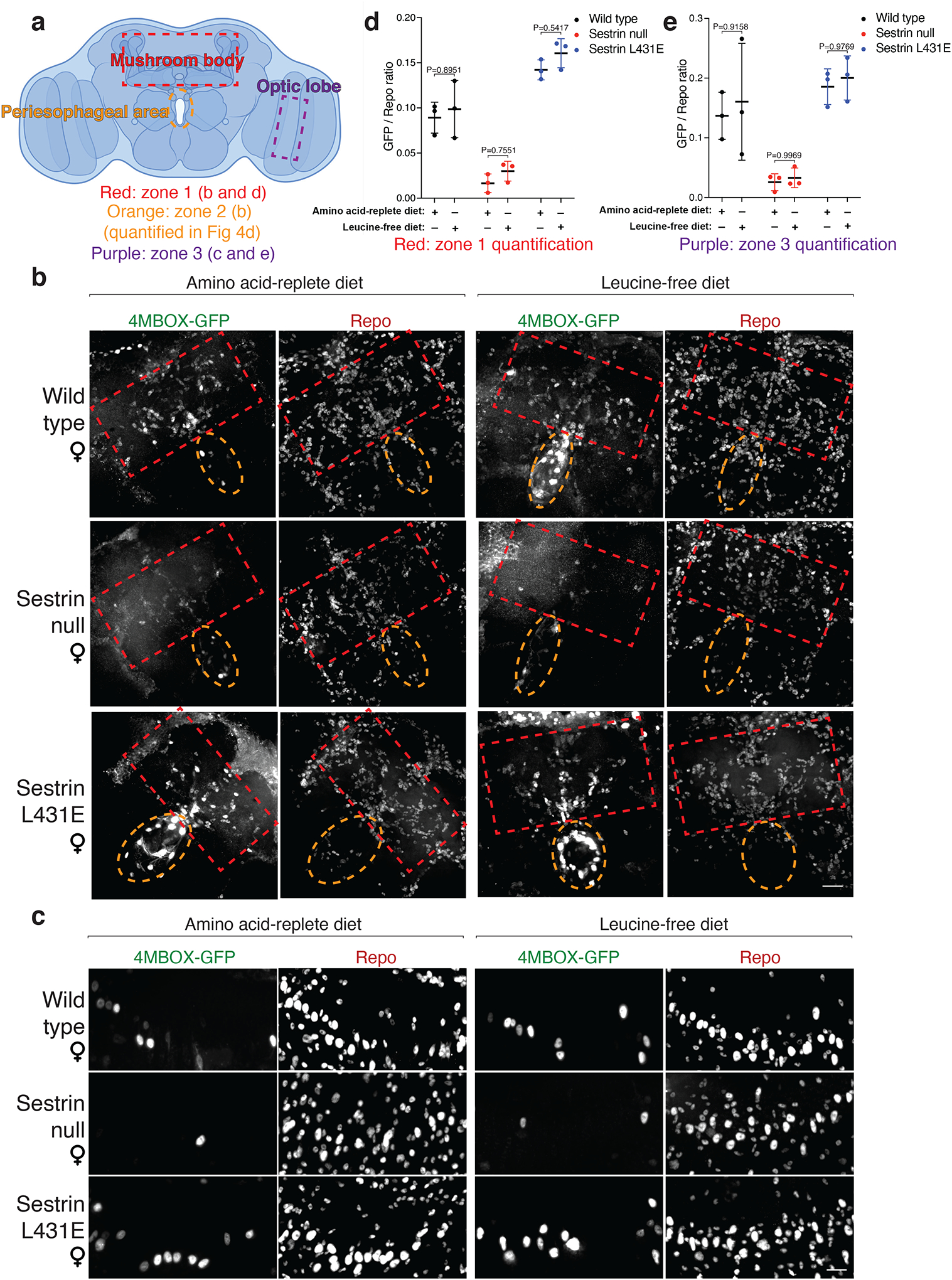
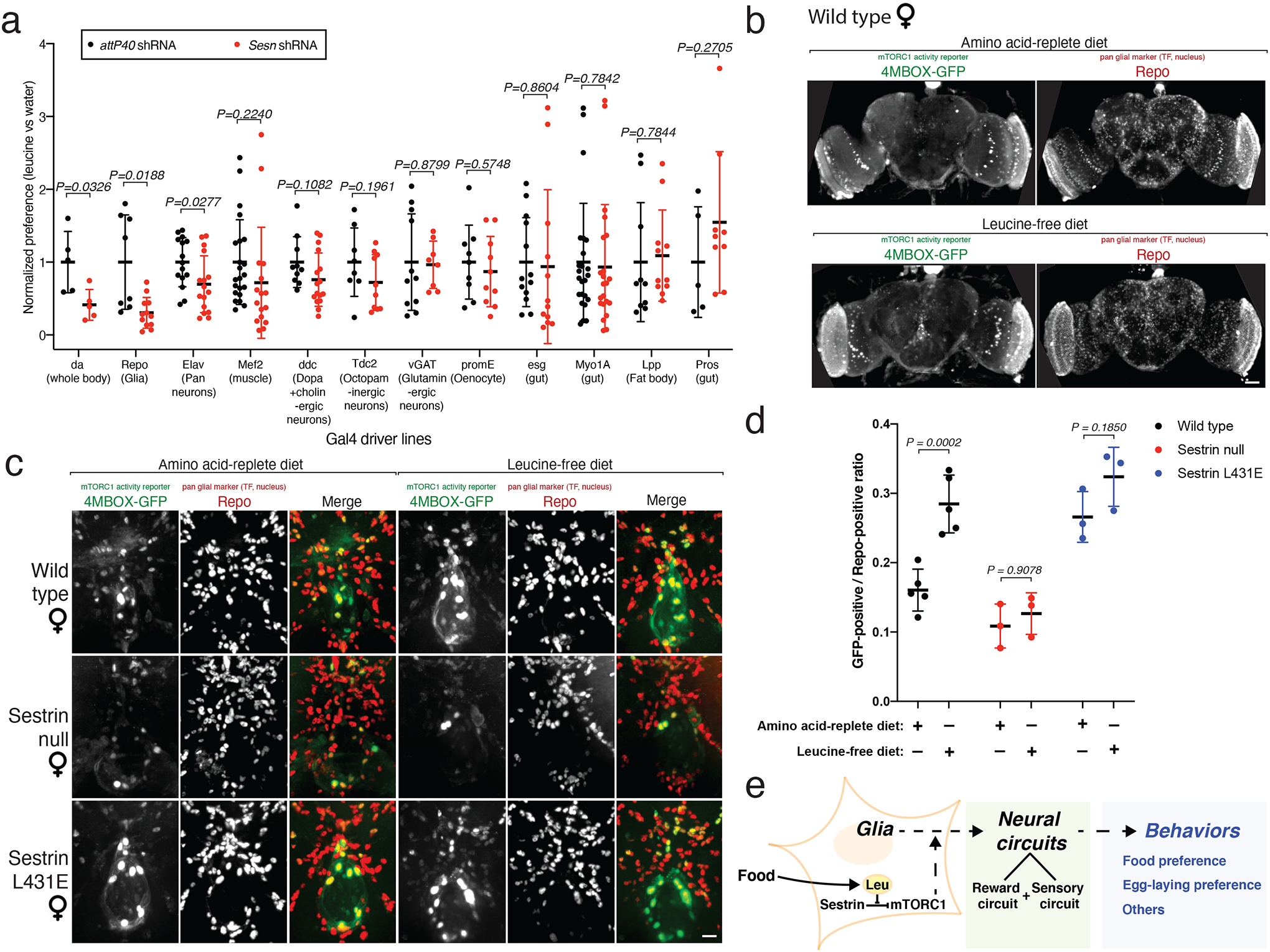
Mechanistic target of rapamycin complex 1 (mTORC1) regulates cell growth and metabolism in response to multiple nutrients, including the essential amino acid leucine1. Recent work in cultured mammalian cells established the Sestrins as leucine-binding proteins that inhibit mTORC1 signalling during leucine deprivation2,3, but their role in the organismal response to dietary leucine remains elusive. Here we find that Sestrin-null flies (Sesn-/-) fail to inhibit mTORC1 or activate autophagy after acute leucine starvation and have impaired development and a shortened lifespan on a low-leucine diet. Knock-in flies expressing a leucine-binding-deficient Sestrin mutant (SesnL431E) have reduced, leucine-insensitive mTORC1 activity. Notably, we find that flies can discriminate between food with or without leucine, and preferentially feed and lay progeny on leucine-containing food. This preference depends on Sestrin and its capacity to bind leucine. Leucine regulates mTORC1 activity in glial cells, and knockdown of Sesn in these cells reduces the ability of flies to detect leucine-free food. Thus, nutrient sensing by mTORC1 is necessary for flies not only to adapt to, but also to detect, a diet deficient in an essential nutrient.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
D.M.S. is a shareholder of Navitor Pharmaceuticals, which is targeting for therapeutic benefit the amino-acid-sensing pathway upstream of mTORC1. J.W.L. advises Raphael Pharmaceuticals, Nanocare Technologies, Petri Biologics, and Restoration Foodworks. M.A.R. is currently employed by Amgen, which has interests in neurodegenerative diseases. These relationships have no overlap with this study. The other authors declare no competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
