

Dynamical landscapes of cell fate decisions
- PMID: 35860004
- PMCID: PMC9184965
- DOI: 10.1098/rsfs.2022.0002
Dynamical landscapes of cell fate decisions
Abstract
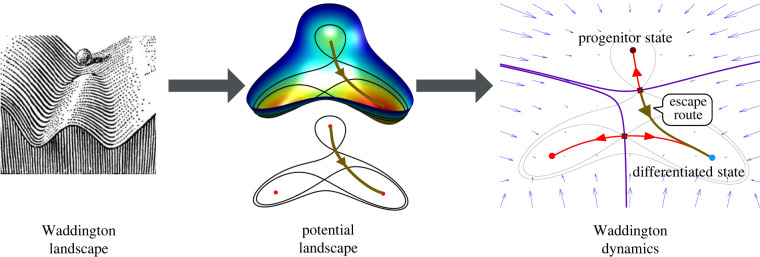
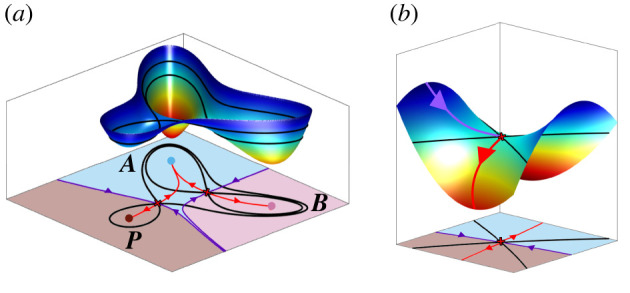
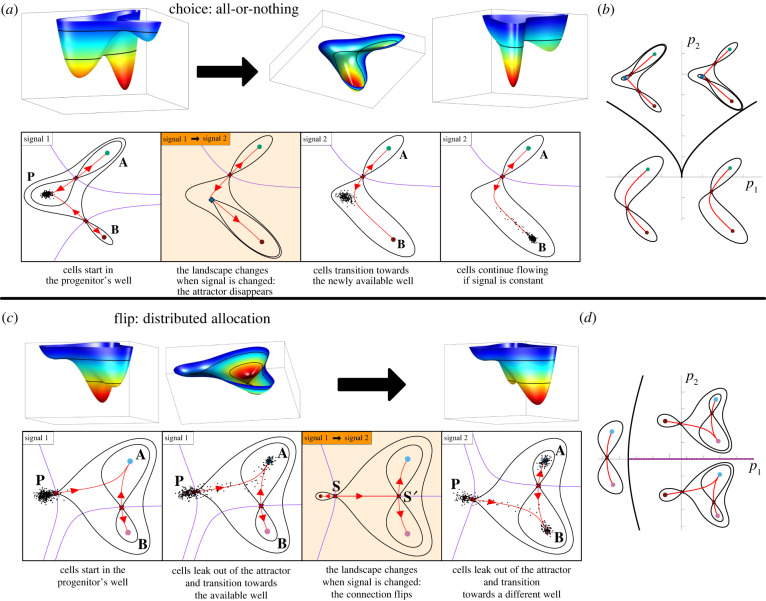
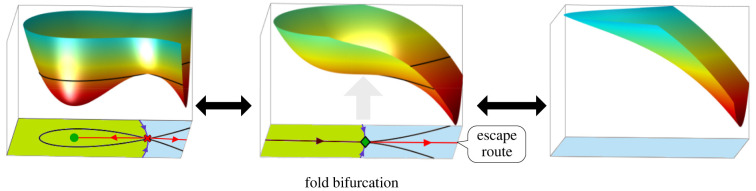
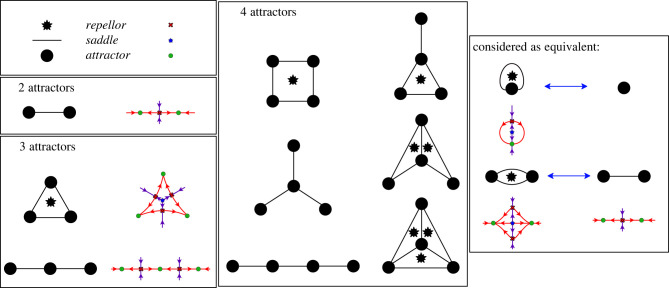
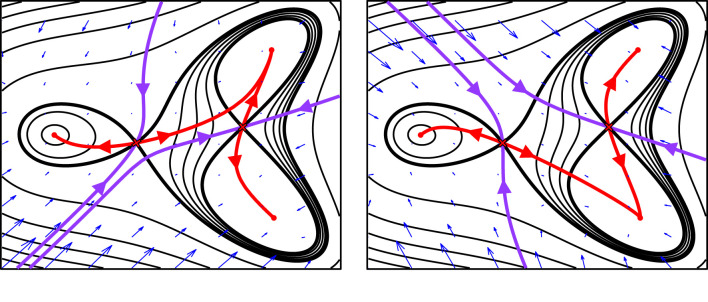
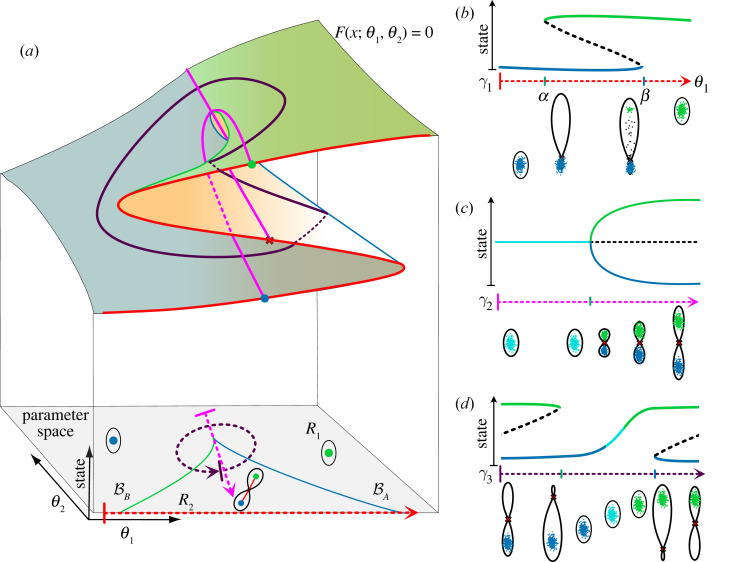
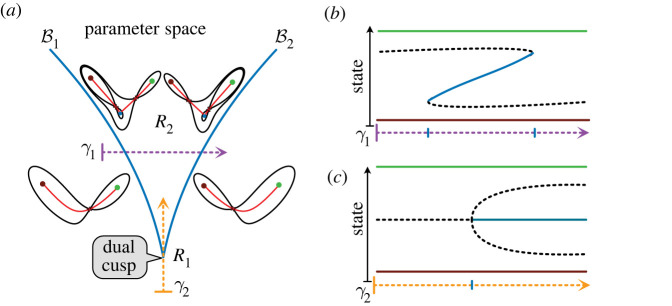
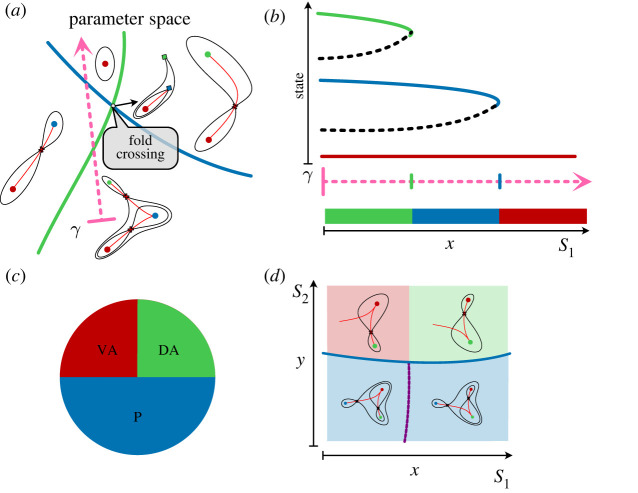
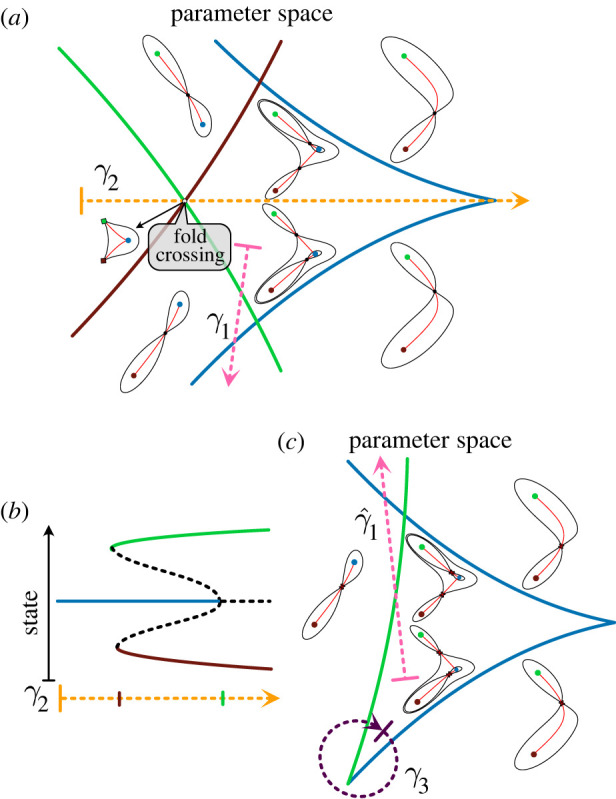
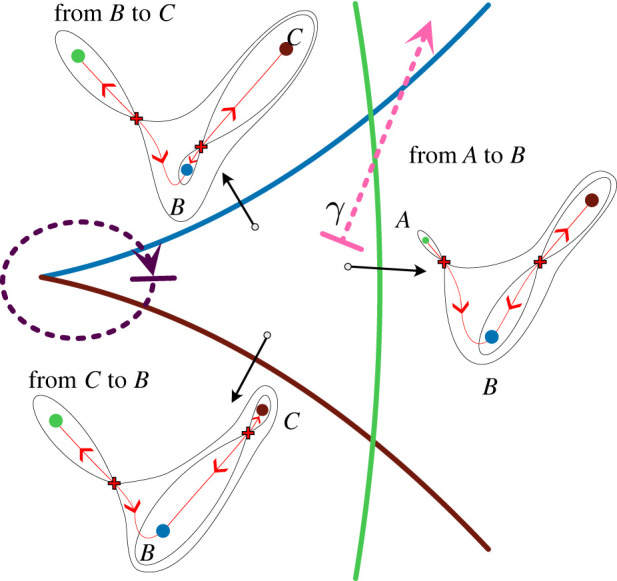
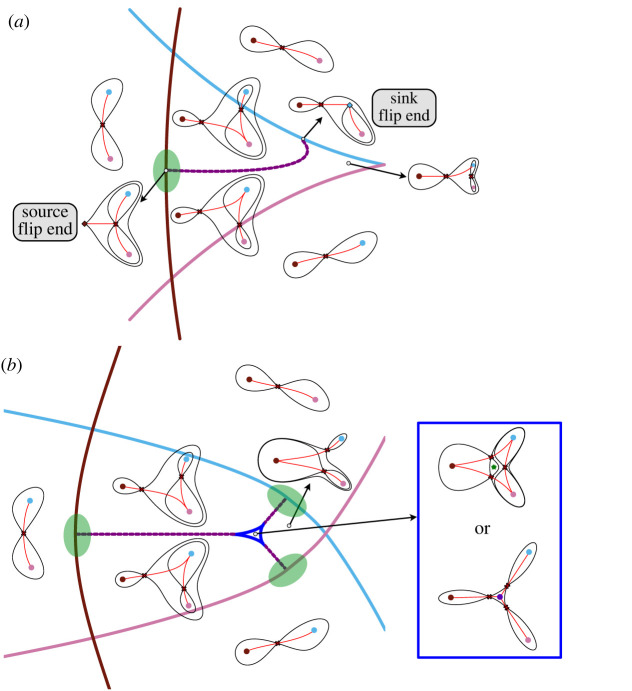
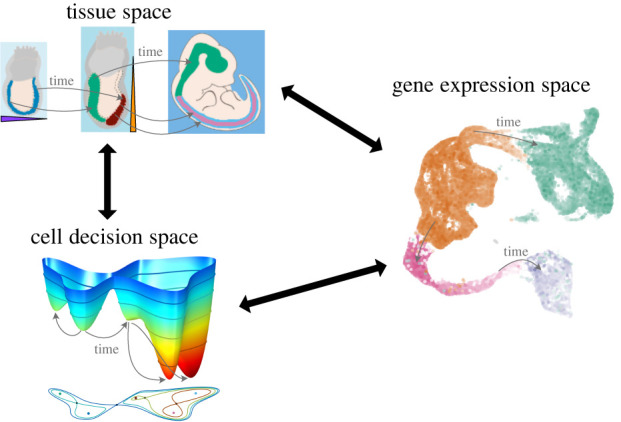
The generation of cellular diversity during development involves differentiating cells transitioning between discrete cell states. In the 1940s, the developmental biologist Conrad Waddington introduced a landscape metaphor to describe this process. The developmental path of a cell was pictured as a ball rolling through a terrain of branching valleys with cell fate decisions represented by the branch points at which the ball decides between one of two available valleys. Here we discuss progress in constructing quantitative dynamical models inspired by this view of cellular differentiation. We describe a framework based on catastrophe theory and dynamical systems methods that provides the foundations for quantitative geometric models of cellular differentiation. These models can be fit to experimental data and used to make quantitative predictions about cellular differentiation. The theory indicates that cell fate decisions can be described by a small number of decision structures, such that there are only two distinct ways in which cells make a binary choice between one of two fates. We discuss the biological relevance of these mechanisms and suggest the approach is broadly applicable for the quantitative analysis of differentiation dynamics and for determining principles of developmental decisions.
Keywords: Waddington landscape; bifurcations; cellular decision-making; development; dynamical systems.
© 2022 The Authors.
Figures
References
-
- Waddington C. 1957. The strategy of the genes. London, UK: Allen & Unwin.
-
- Smale S. 1967. Differentiable dynamical systems. Bull. Am. Math. Soc. 73, 747-817. ( 10.1090/S0002-9904-1967-11798-1) - DOI
-
- Thom R. 1989. Structural stability and morphogenesis: an outline of a general theory of models. Boca Raton, FL: CRC Press.
-
- Zeeman EC. 1977. Catastrophe theory: selected papers, 1972–1977. Advanced Book Program. Reading, MA: Addison-Wesley.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources