

Telomere Length Regulation
- PMID: 35860550
- PMCID: PMC9289283
- DOI: 10.3389/fonc.2022.943622
Telomere Length Regulation
Abstract
The number of (TTAGGG)n repeats at the ends of chromosomes is highly variable between individual chromosomes, between different cells and between species. Progressive loss of telomere repeats limits the proliferation of pre-malignant human cells but also contributes to aging by inducing apoptosis and senescence in normal cells. Despite enormous progress in understanding distinct pathways that result in loss and gain of telomeric DNA in different cell types, many questions remain. Further studies are needed to delineate the role of damage to telomeric DNA, replication errors, chromatin structure, liquid-liquid phase transition, telomeric transcripts (TERRA) and secondary DNA structures such as guanine quadruplex structures, R-loops and T-loops in inducing gains and losses of telomere repeats in different cell types. Limitations of current telomere length measurements techniques and differences in telomere biology between species and different cell types complicate generalizations about the role of telomeres in aging and cancer. Here some of the factors regulating the telomere length in embryonic and adult cells in mammals are discussed from a mechanistic and evolutionary perspective.
Keywords: development; lifespan; quadruplex DNA; telomerase; telomere length measurements; telomere length regulation; telomere replication; tumor suppression.
Copyright © 2022 Lansdorp.
Conflict of interest statement
The author is a founder and shareholder of Repeat Diagnostics Inc., a company specializing in clinical telomere length measurements.
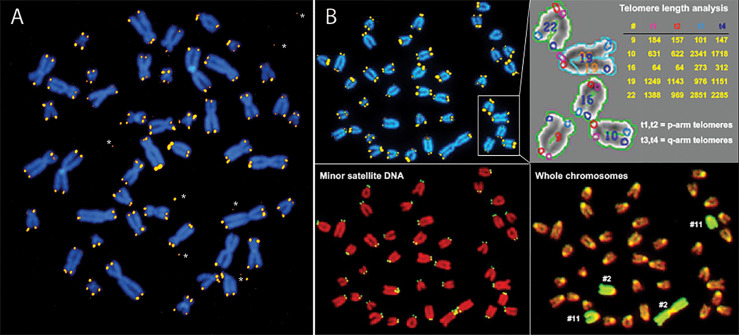
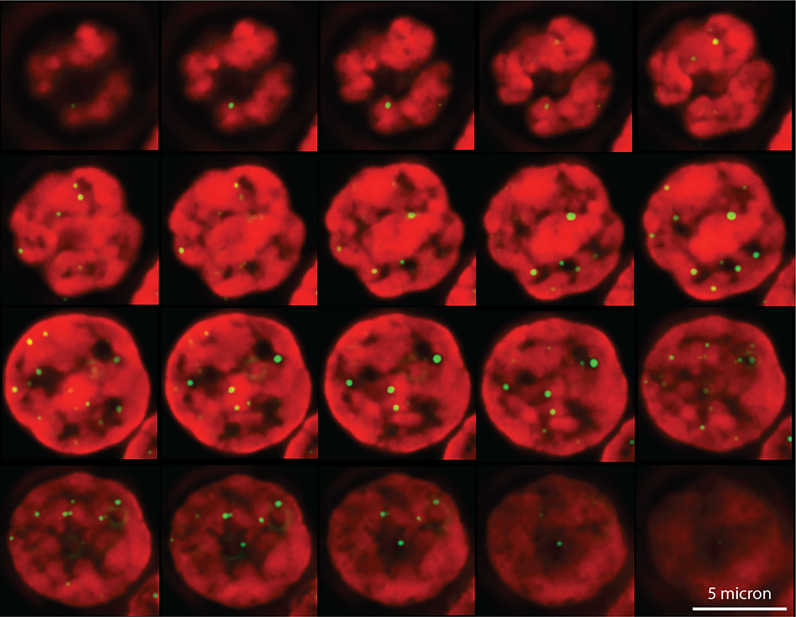
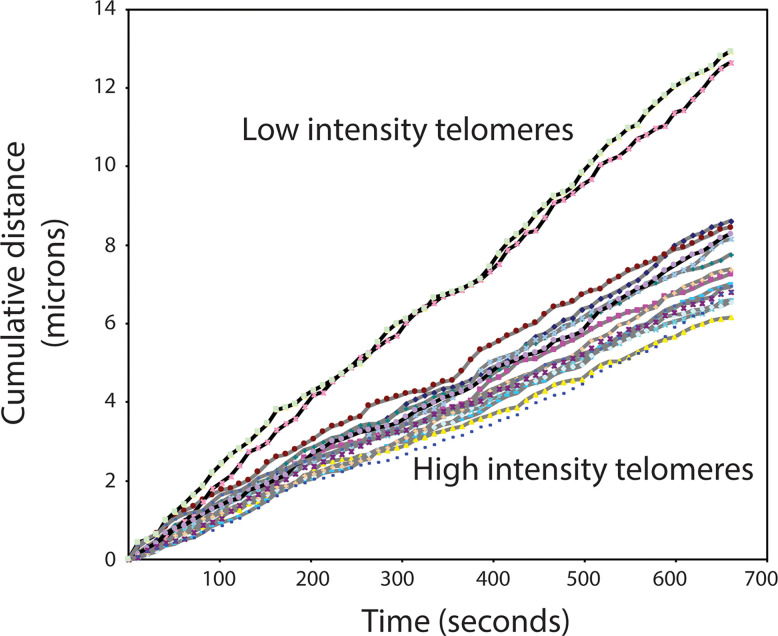
Figures
References
Publication types
LinkOut - more resources
Full Text Sources