

Transcription factors ZmNF-YA1 and ZmNF-YB16 regulate plant growth and drought tolerance in maize
- PMID: 35861438
- PMCID: PMC9516732
- DOI: 10.1093/plphys/kiac340
Transcription factors ZmNF-YA1 and ZmNF-YB16 regulate plant growth and drought tolerance in maize
Abstract
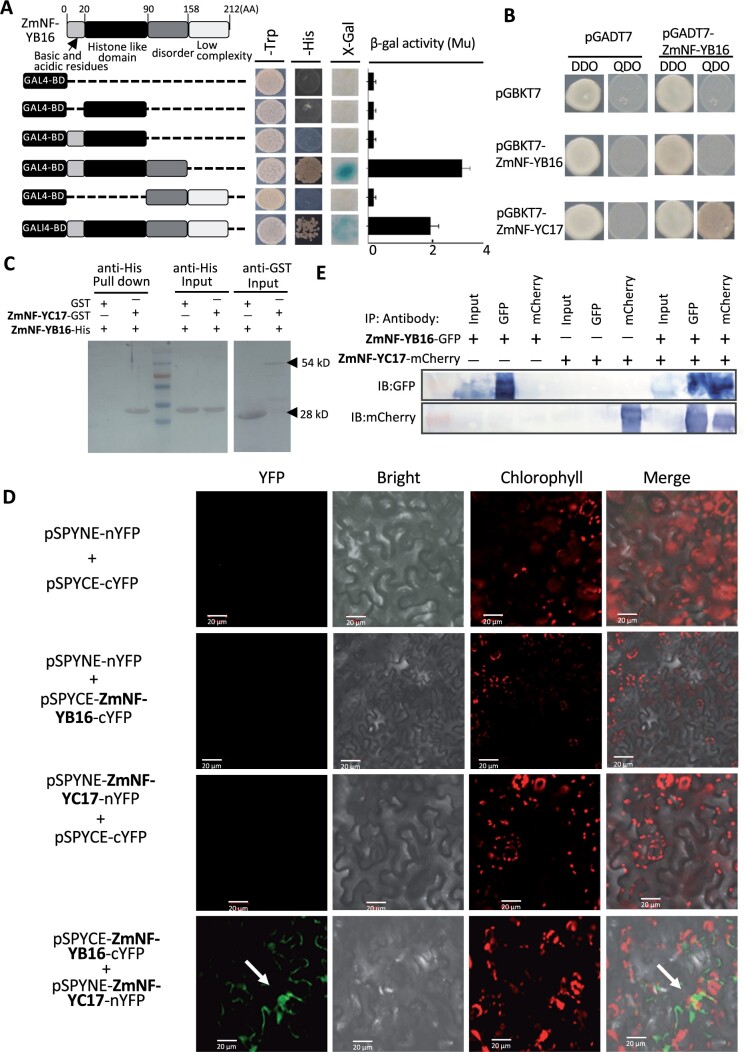
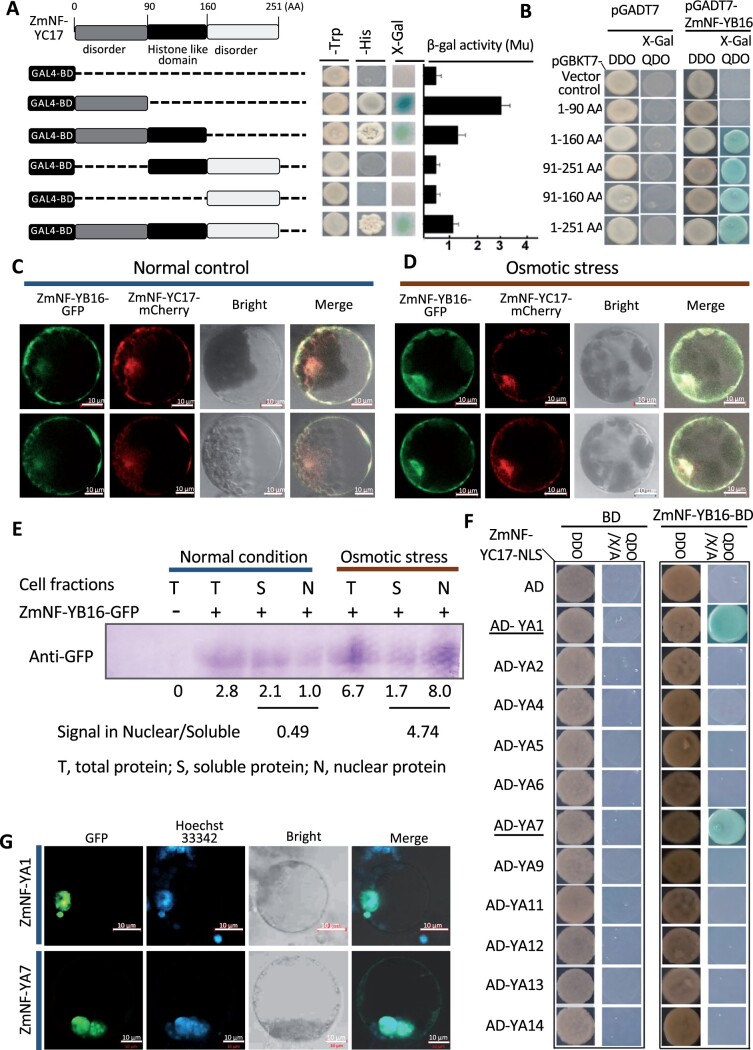
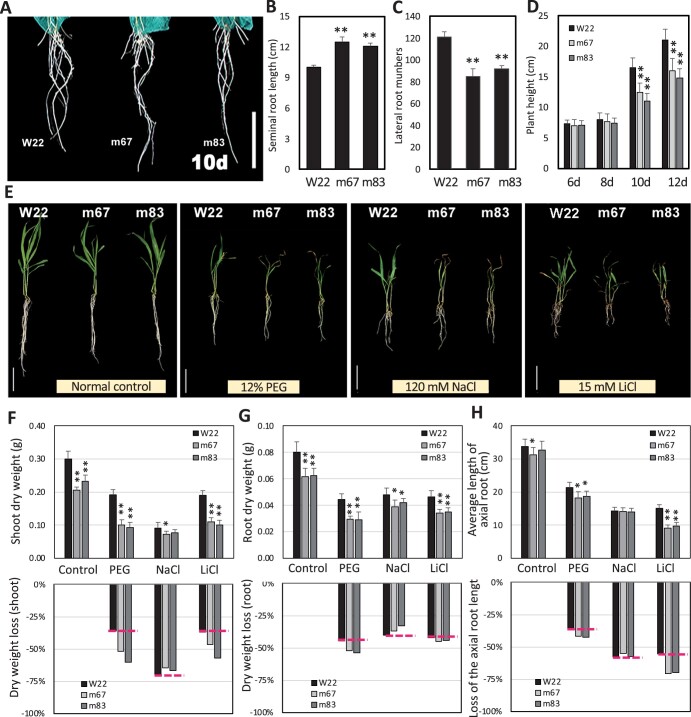
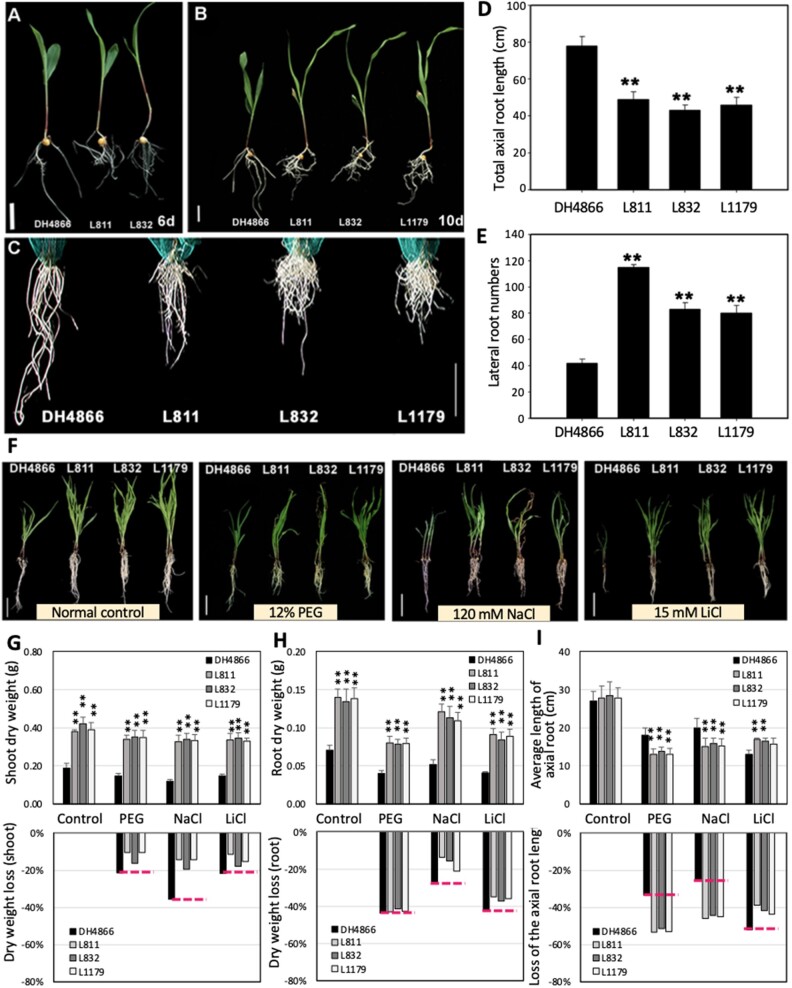
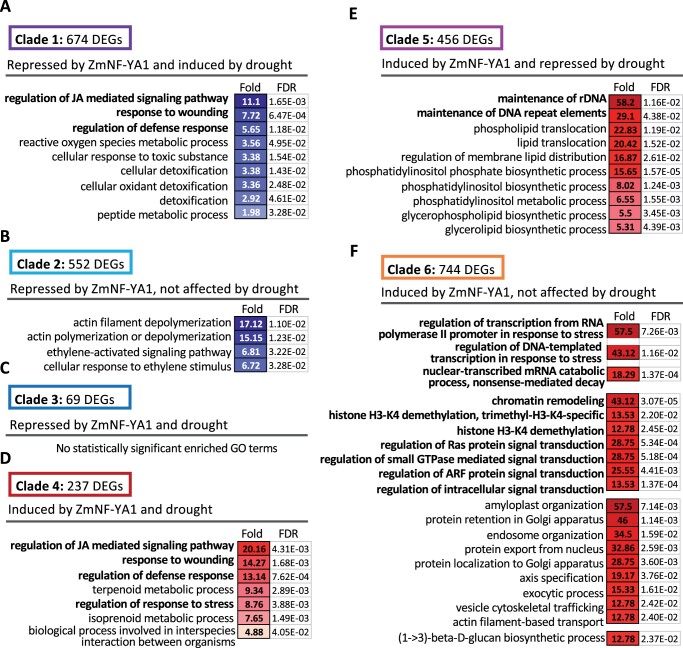
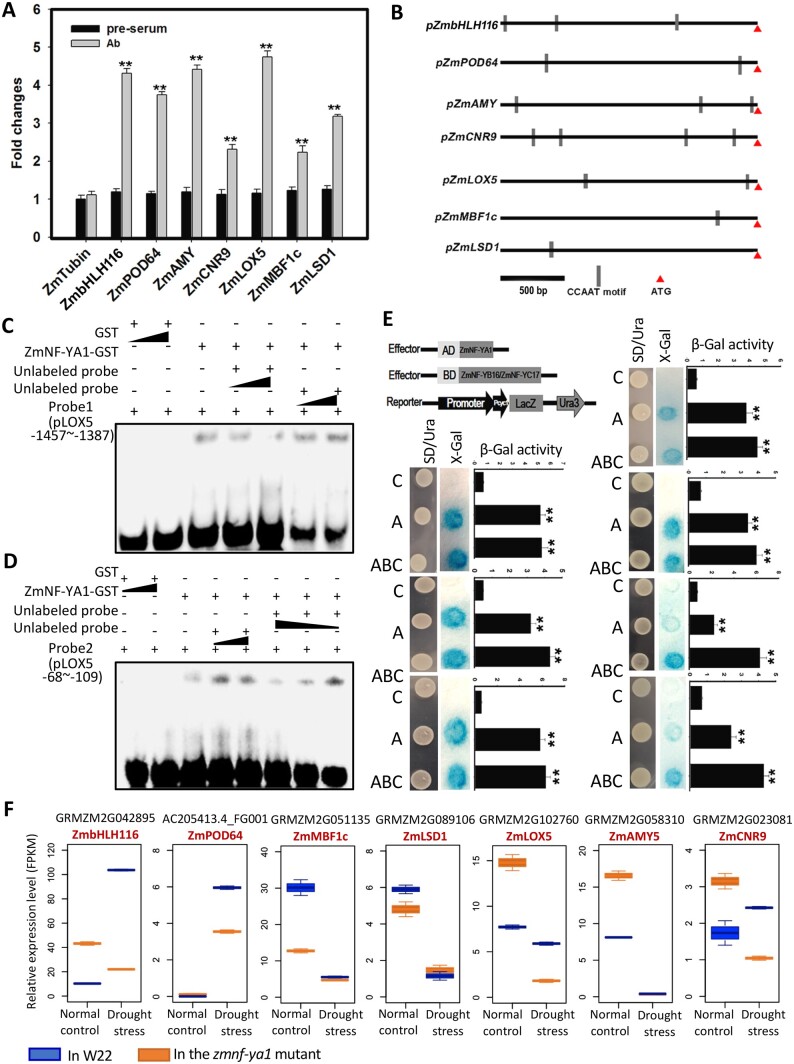
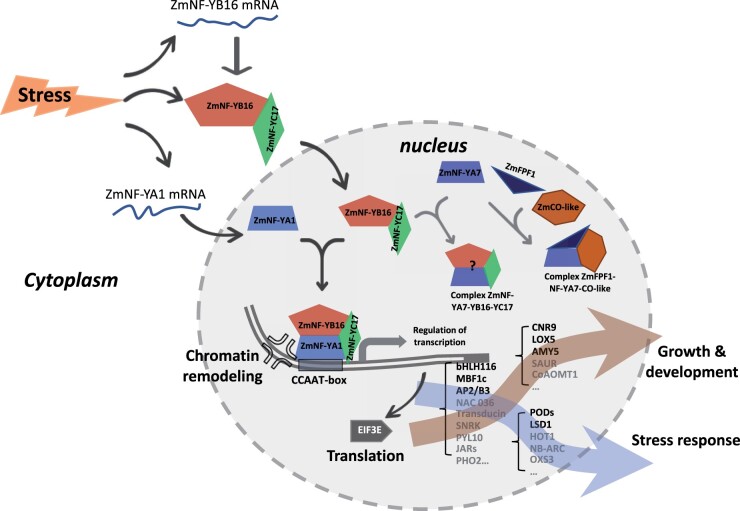
The identification of drought stress regulatory genes is crucial for the genetic improvement of maize (Zea mays L.) yield. Nuclear factors Y (NF-Ys) are important transcription factors, but their roles in the drought stress tolerance of plants and underlying molecular mechanisms are largely unknown. In this work, we used yeast two-hybrid screening to identify potential interactors of ZmNF-YB16 and confirmed the interaction between ZmNF-YA1 and ZmNF-YB16-YC17 and between ZmNF-YA7 and ZmNF-YB16-YC17. ZmNF-YB16 interacted with ZmNF-YC17 via its histone fold domain to form a heterodimer in the cytoplasm and then entered the nucleus to form a heterotrimer with ZmNF-YA1 or ZmNF-YA7 under osmotic stress. Overexpression of ZmNF-YA1 improved drought and salt stress tolerance and root development of maize, whereas zmnf-ya1 mutants exhibited drought and salt stress sensitivity. ZmNF-YA1-mediated transcriptional regulation, especially in JA signaling, histone modification, and chromatin remodeling, could underlie the altered stress tolerance of zmnf-ya1 mutant plants. ZmNF-YA1 bound to promoter CCAAT motifs and directly regulated the expression of multiple genes that play important roles in stress responses and plant development. Comparison of ZmNF-YB16- and ZmNF-YA1-regulated genes showed that ZmNF-YA1 and ZmNF-YB16 have similar biological functions in stress responses but varied functions in other biological processes. Taken together, ZmNF-YA1 is a positive regulator of plant drought and salt stress responses and is involved in the root development of maize, and ZmNF-Y complexes with different subunits may have discrepant functions.
© The Author(s) 2022. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures
Similar articles
-
Genetic, molecular and physiological crosstalk during drought tolerance in maize (Zea mays): pathways to resilient agriculture.Planta. 2024 Aug 28;260(4):81. doi: 10.1007/s00425-024-04517-9. Planta. 2024. PMID: 39196449 Review.
-
ZmNF-YB10, a maize NF-Y transcription factor, positively regulates drought and salt stress response in Arabidopsis thaliana.GM Crops Food. 2025 Dec;16(1):28-45. doi: 10.1080/21645698.2024.2438421. Epub 2024 Dec 24. GM Crops Food. 2025. PMID: 39718137 Free PMC article.
-
ZmNF-YA1 Contributes to Maize Thermotolerance by Regulating Heat Shock Response.Int J Mol Sci. 2024 Jun 6;25(11):6275. doi: 10.3390/ijms25116275. Int J Mol Sci. 2024. PMID: 38892463 Free PMC article.
-
Overexpression of maize transcription factor ZmNF-YC14 positively regulates drought and salt stress responses in Arabidopsis thaliana.Plant Sci. 2025 Jul;356:112502. doi: 10.1016/j.plantsci.2025.112502. Epub 2025 Apr 7. Plant Sci. 2025. PMID: 40204192
-
Transcription factors as molecular switches to regulate drought adaptation in maize.Theor Appl Genet. 2020 May;133(5):1455-1465. doi: 10.1007/s00122-019-03494-y. Epub 2019 Dec 5. Theor Appl Genet. 2020. PMID: 31807836 Review.
Cited by
-
Drought stress memory in maize: understanding and harnessing the past for future resilience.Plant Cell Rep. 2025 Apr 25;44(5):101. doi: 10.1007/s00299-025-03494-x. Plant Cell Rep. 2025. PMID: 40278890 Review.
-
Genetic, molecular and physiological crosstalk during drought tolerance in maize (Zea mays): pathways to resilient agriculture.Planta. 2024 Aug 28;260(4):81. doi: 10.1007/s00425-024-04517-9. Planta. 2024. PMID: 39196449 Review.
-
Crucial Abiotic Stress Regulatory Network of NF-Y Transcription Factor in Plants.Int J Mol Sci. 2023 Feb 23;24(5):4426. doi: 10.3390/ijms24054426. Int J Mol Sci. 2023. PMID: 36901852 Free PMC article. Review.
-
Recent advances in improving yield and immunity through transcription factor engineering.J Integr Plant Biol. 2025 Aug;67(8):2005-2027. doi: 10.1111/jipb.13932. Epub 2025 May 21. J Integr Plant Biol. 2025. PMID: 40396540 Free PMC article. Review.
-
ZmNF-YB10, a maize NF-Y transcription factor, positively regulates drought and salt stress response in Arabidopsis thaliana.GM Crops Food. 2025 Dec;16(1):28-45. doi: 10.1080/21645698.2024.2438421. Epub 2024 Dec 24. GM Crops Food. 2025. PMID: 39718137 Free PMC article.
References
-
- Alam MM, Tanaka T, Nakamura H, Ichikawa H, Kobayashi K, Yaeno T, Yamaoka N, Shimomoto K, Takayama K, Nishina H, et al. (2015) Overexpression of a rice heme activator protein gene (OsHAP2E) confers resistance to pathogens, salinity and drought, and increases photosynthesis and tiller number. Plant Biotechnol J 13: 85–96 - PubMed
-
- Ballif J, Endo S, Kotani M, MacAdam J, Wu Y (2011) Over-expression of HAP3b enhances primary root elongation in Arabidopsis. Plant Physiol Biochem 49: 579–583 - PubMed
-
- Frontini M, Imbriano C, Manni I, Mantovani R (2004) Cell cycle regulation of NF-YC nuclear localization. Cell Cycle 3: 217–222 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
