

Cardiomyocyte-Specific Long Noncoding RNA Regulates Alternative Splicing of the Triadin Gene in the Heart
- PMID: 35862102
- PMCID: PMC9427731
- DOI: 10.1161/CIRCULATIONAHA.121.058017
Cardiomyocyte-Specific Long Noncoding RNA Regulates Alternative Splicing of the Triadin Gene in the Heart
Abstract
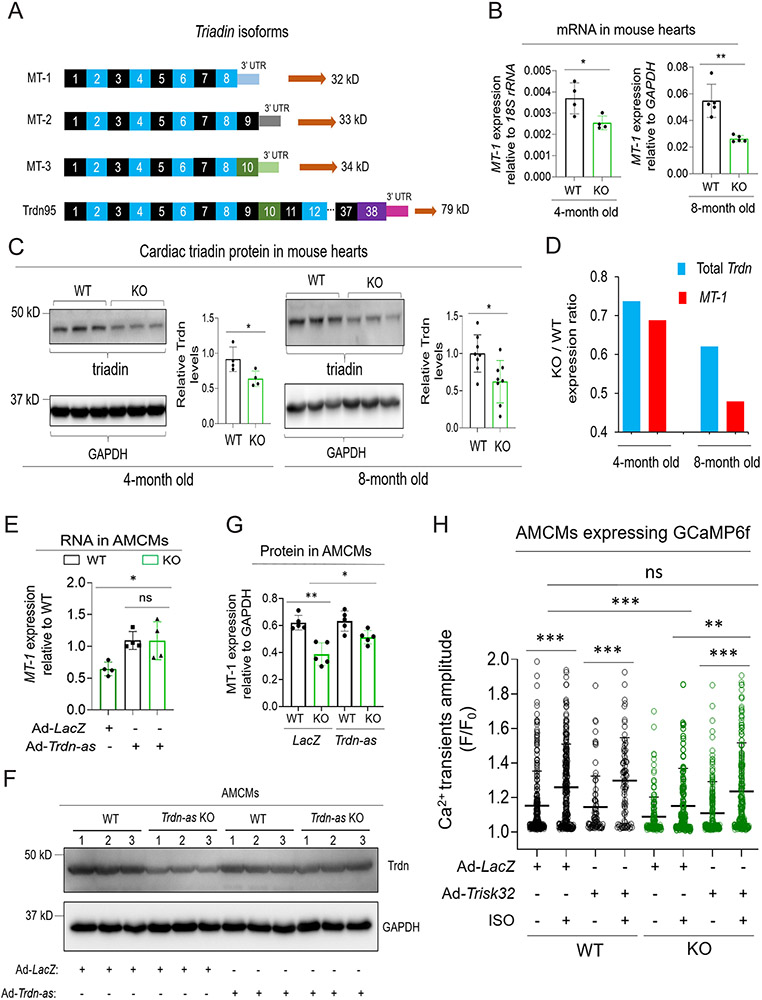
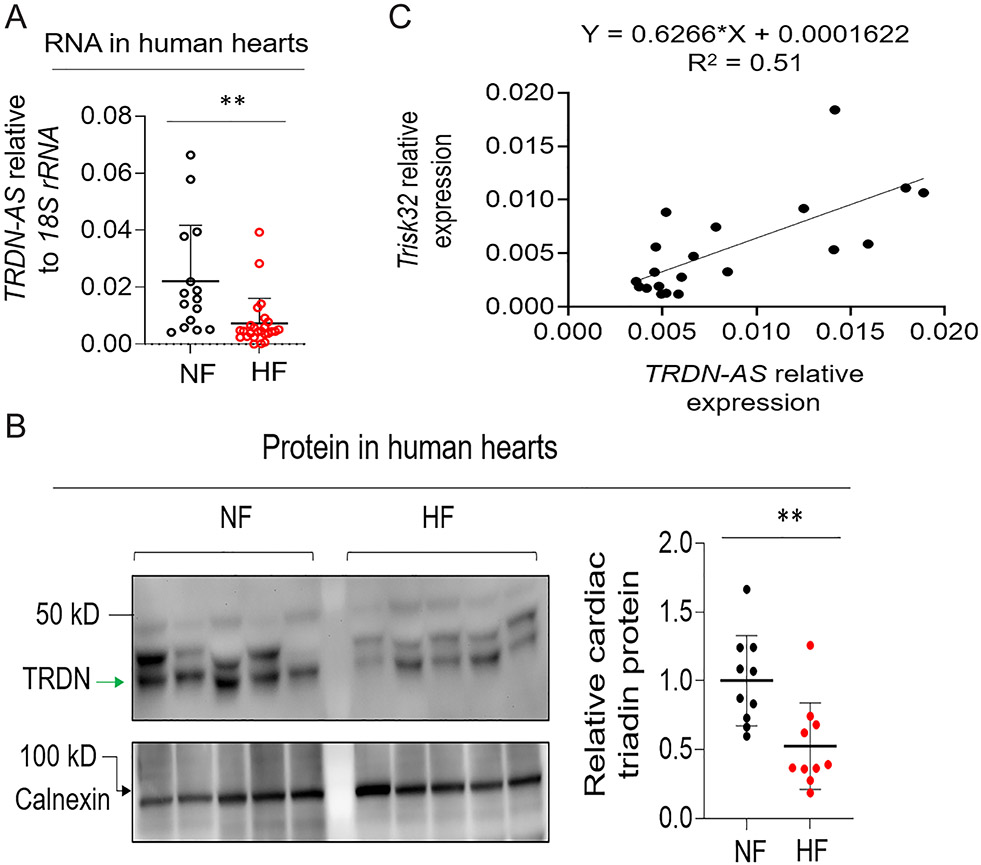
Background: Abnormalities in Ca2+ homeostasis are associated with cardiac arrhythmias and heart failure. Triadin plays an important role in Ca2+ homeostasis in cardiomyocytes. Alternative splicing of a single triadin gene produces multiple triadin isoforms. The cardiac-predominant isoform, mouse MT-1 or human Trisk32, is encoded by triadin exons 1 to 8. In humans, mutations in the triadin gene that lead to a reduction in Trisk32 levels in the heart can cause cardiac dysfunction and arrhythmias. Decreased levels of Trisk32 in the heart are also common in patients with heart failure. However, mechanisms that maintain triadin isoform composition in the heart remain elusive.
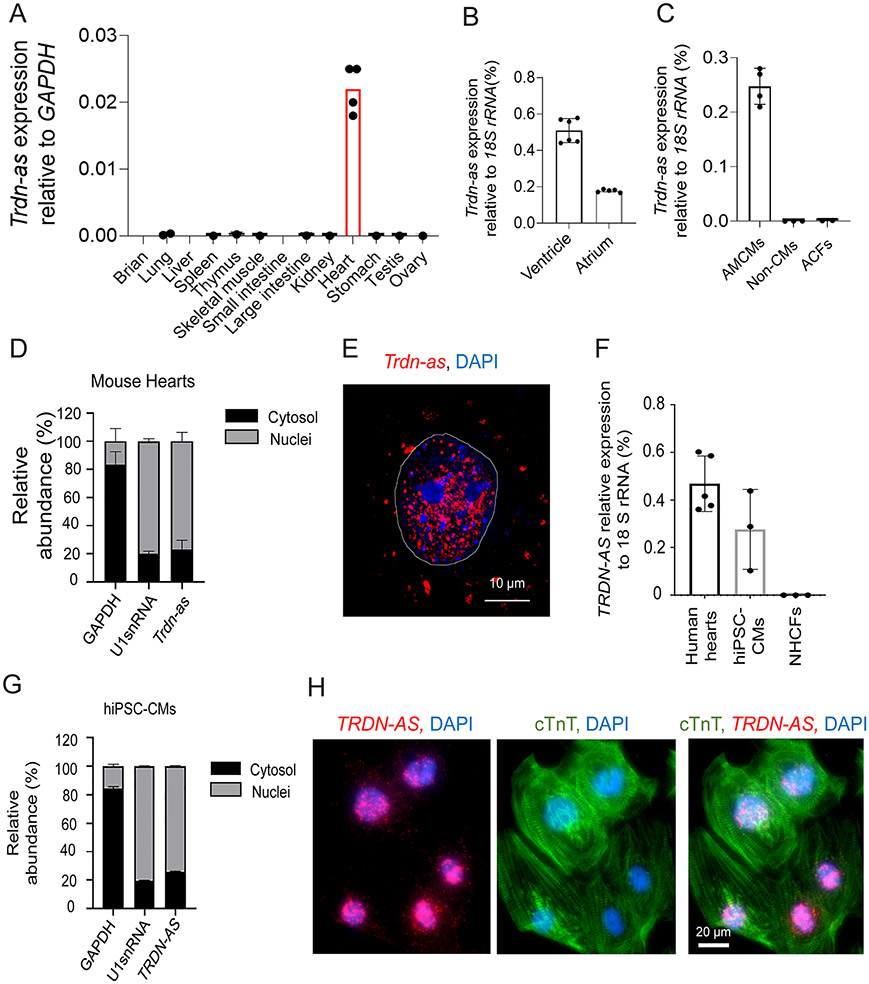
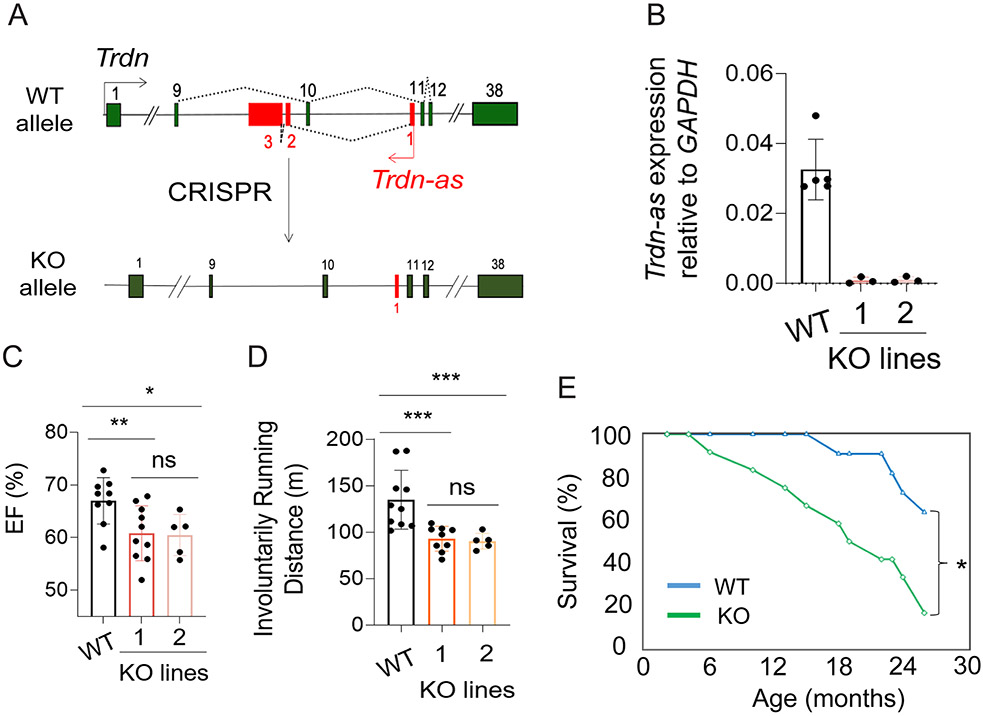
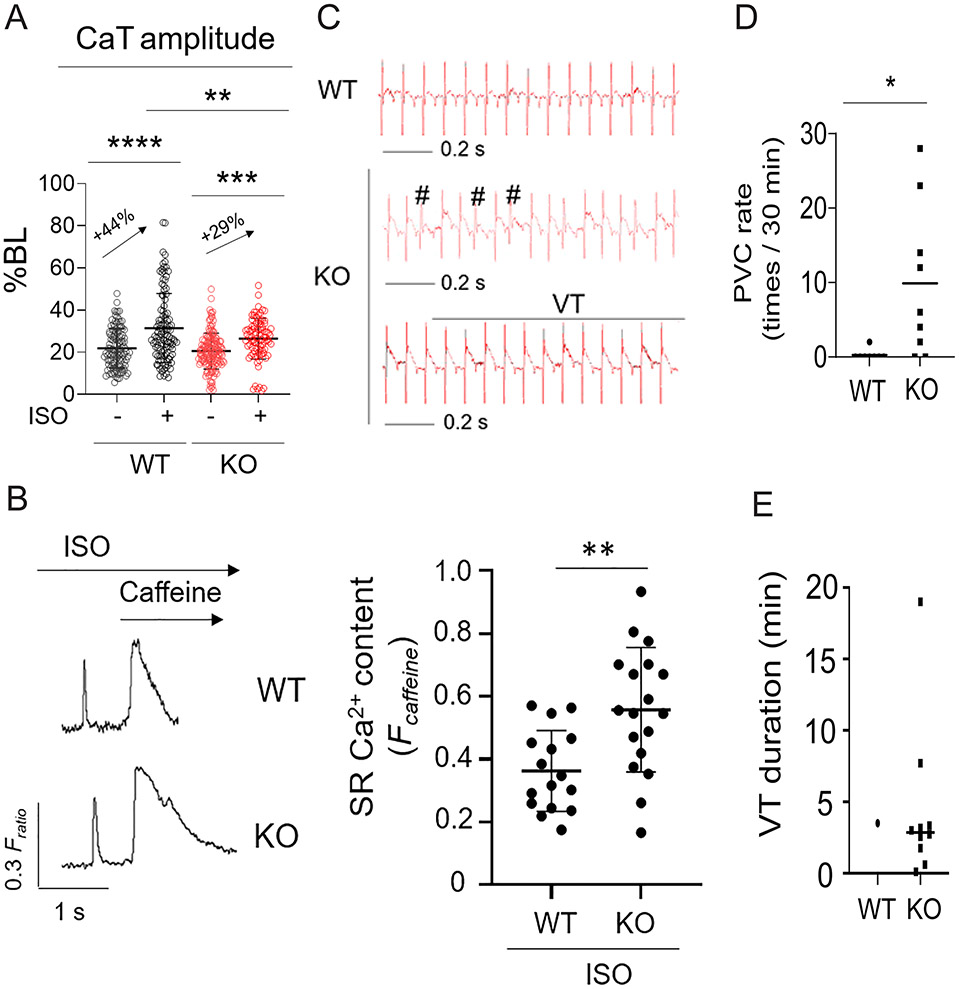
Methods: We analyzed triadin expression in heart explants from patients with heart failure and cardiac arrhythmias and in hearts from mice carrying a knockout allele for Trdn-as, a cardiomyocyte-specific long noncoding RNA encoded by the antisense strand of the triadin gene, between exons 9 and 11. Catecholamine challenge with isoproterenol was performed on Trdn-as knockout mice to assess the role of Trdn-as in cardiac arrhythmogenesis, as assessed by ECG. Ca2+ transients in adult mouse cardiomyocytes were measured with the IonOptix platform or the GCaMP system. Biochemistry assays, single-molecule fluorescence in situ hybridization, subcellular localization imaging, RNA sequencing, and molecular rescue assays were used to investigate the mechanisms by which Trdn-as regulates cardiac function and triadin levels in the heart.
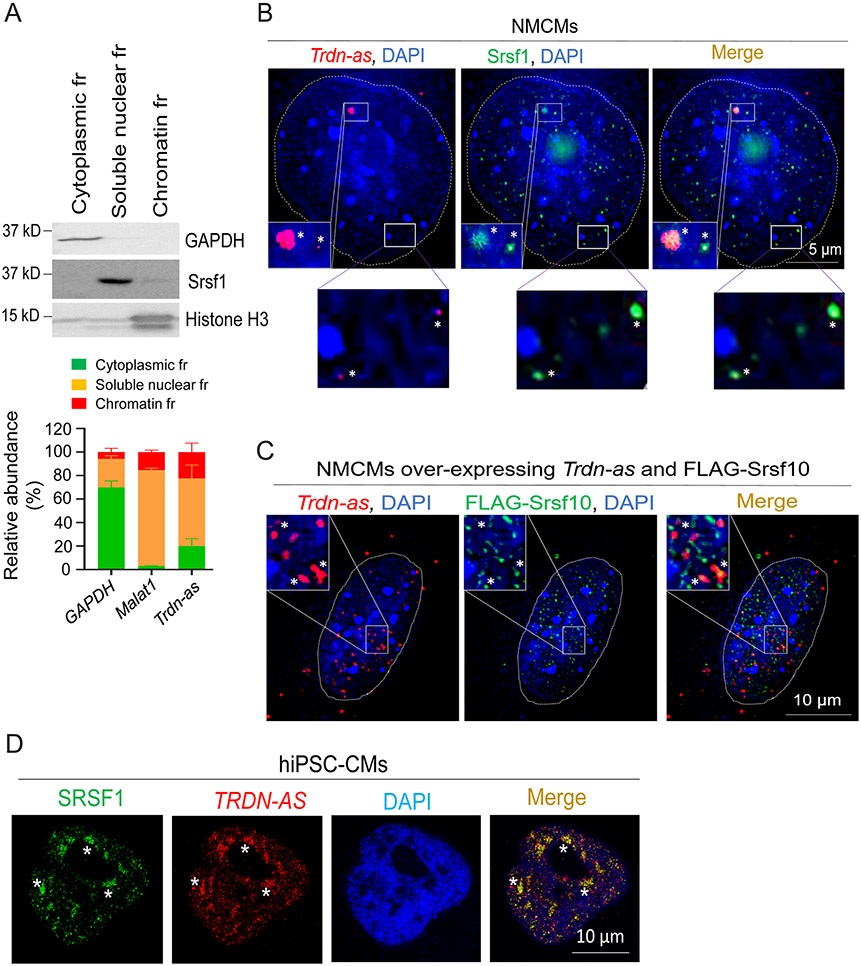
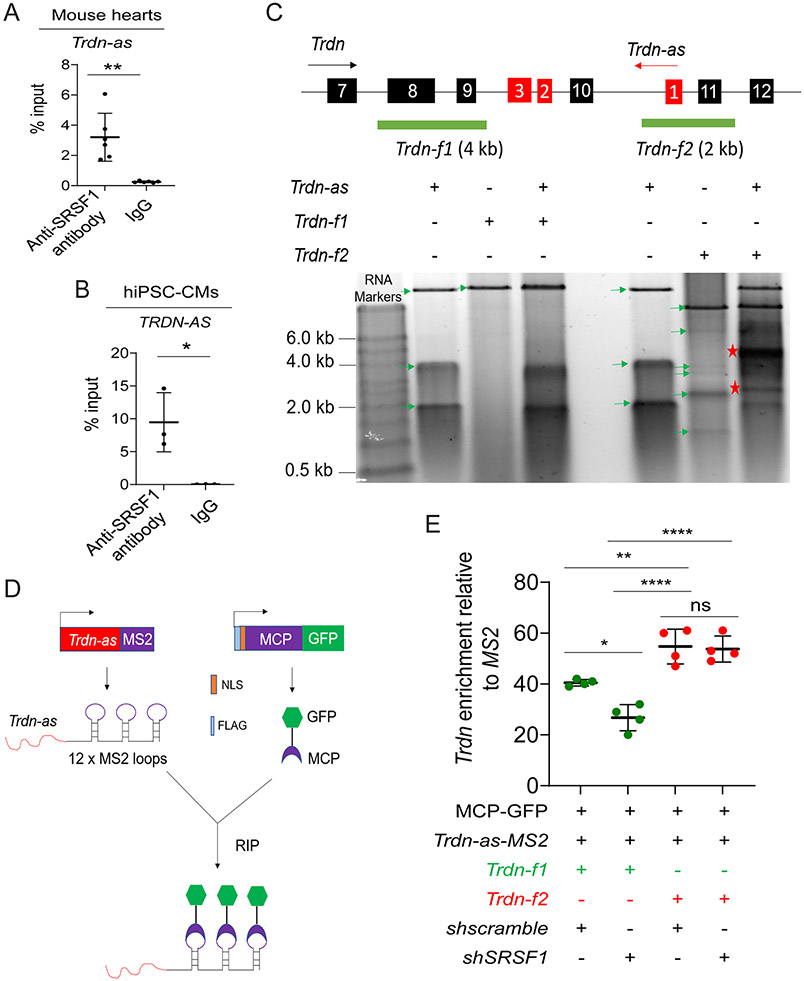
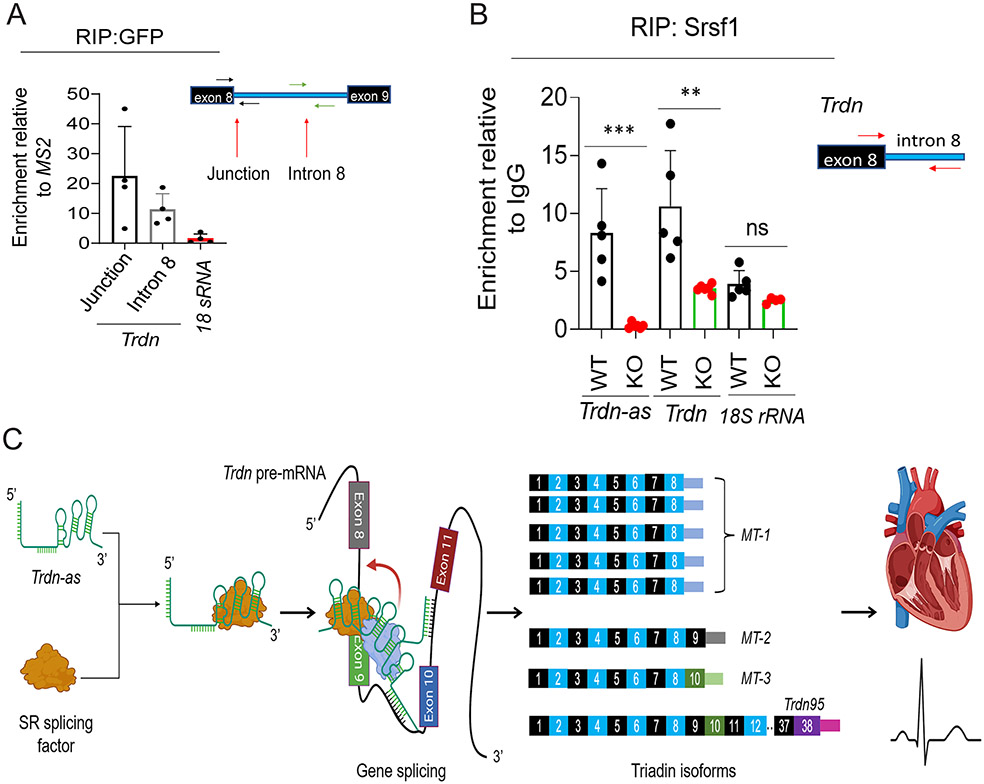
Results: We report that Trdn-as maintains cardiac function, at least in part, by regulating alternative splicing of the triadin gene. Knockout of Trdn-as in mice downregulates cardiac triadin, impairs Ca2+ handling, and causes premature death. Trdn-as knockout mice are susceptible to cardiac arrhythmias in response to catecholamine challenge. Normalization of cardiac triadin levels in Trdn-as knockout cardiomyocytes is sufficient to restore Ca2+ handling. Last, Trdn-as colocalizes and interacts with serine/arginine splicing factors in cardiomyocyte nuclei and is essential for efficient recruitment of splicing factors to triadin precursor mRNA.
Conclusions: These findings reveal regulation of alternative splicing as a novel mechanism by which a long noncoding RNA controls cardiac function. This study indicates potential therapeutics for heart disease by targeting the long noncoding RNA or pathways regulating alternative splicing.
Keywords: RNA, long noncoding; arrhythmias, cardiac; heart failure; myocytes, cardiac; splicing, alternative.
Figures
Comment in
-
Triadin-Antisense: An lncRNA in the Backstage of Cardiac Alternative Splicing.Circulation. 2022 Aug 30;146(9):715-717. doi: 10.1161/CIRCULATIONAHA.122.061232. Epub 2022 Aug 29. Circulation. 2022. PMID: 36037268 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
