

Chitin-Active Lytic Polysaccharide Monooxygenases Are Rare in Cellulomonas Species
- PMID: 35862679
- PMCID: PMC9361826
- DOI: 10.1128/aem.00968-22
Chitin-Active Lytic Polysaccharide Monooxygenases Are Rare in Cellulomonas Species
Abstract
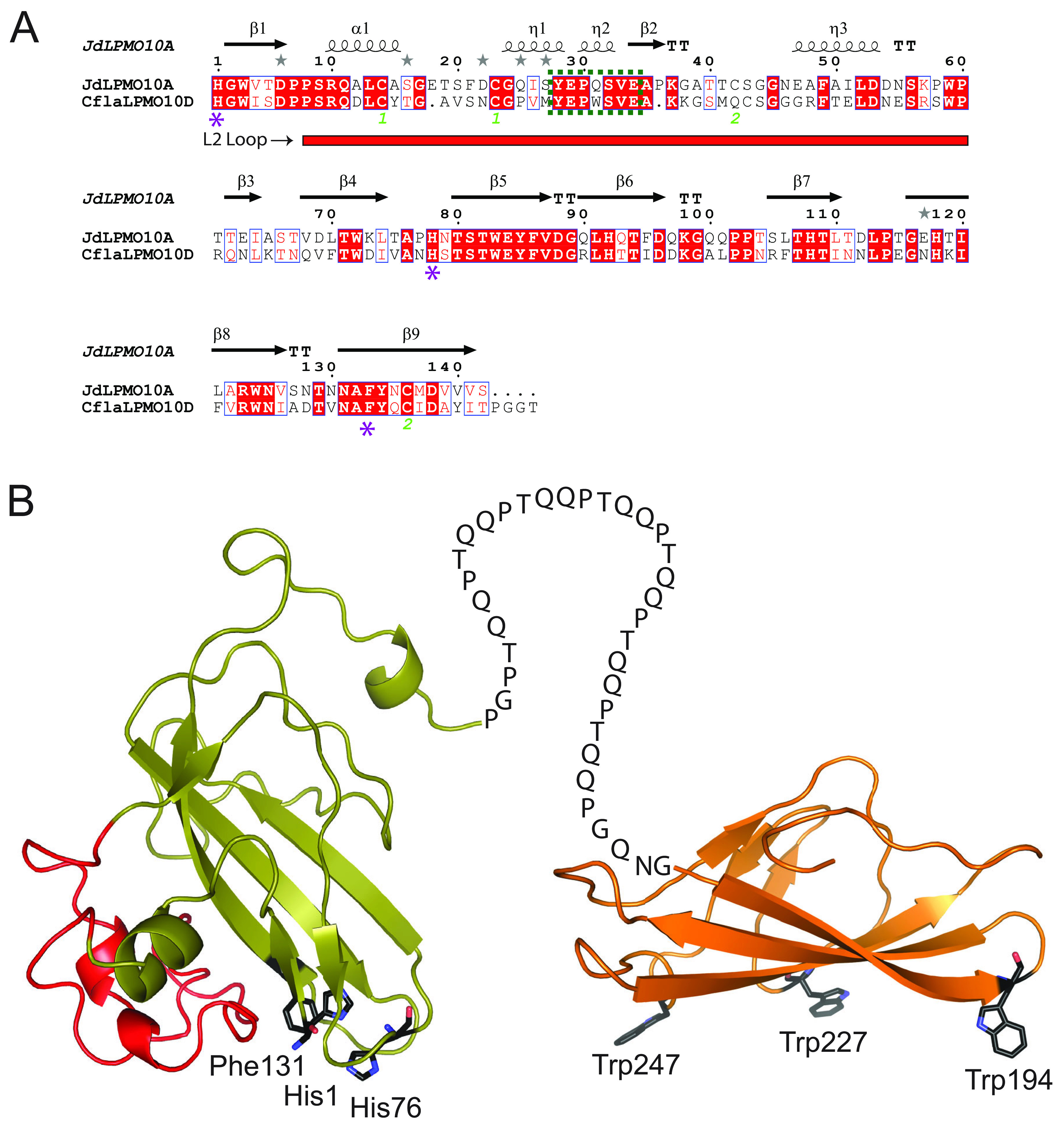
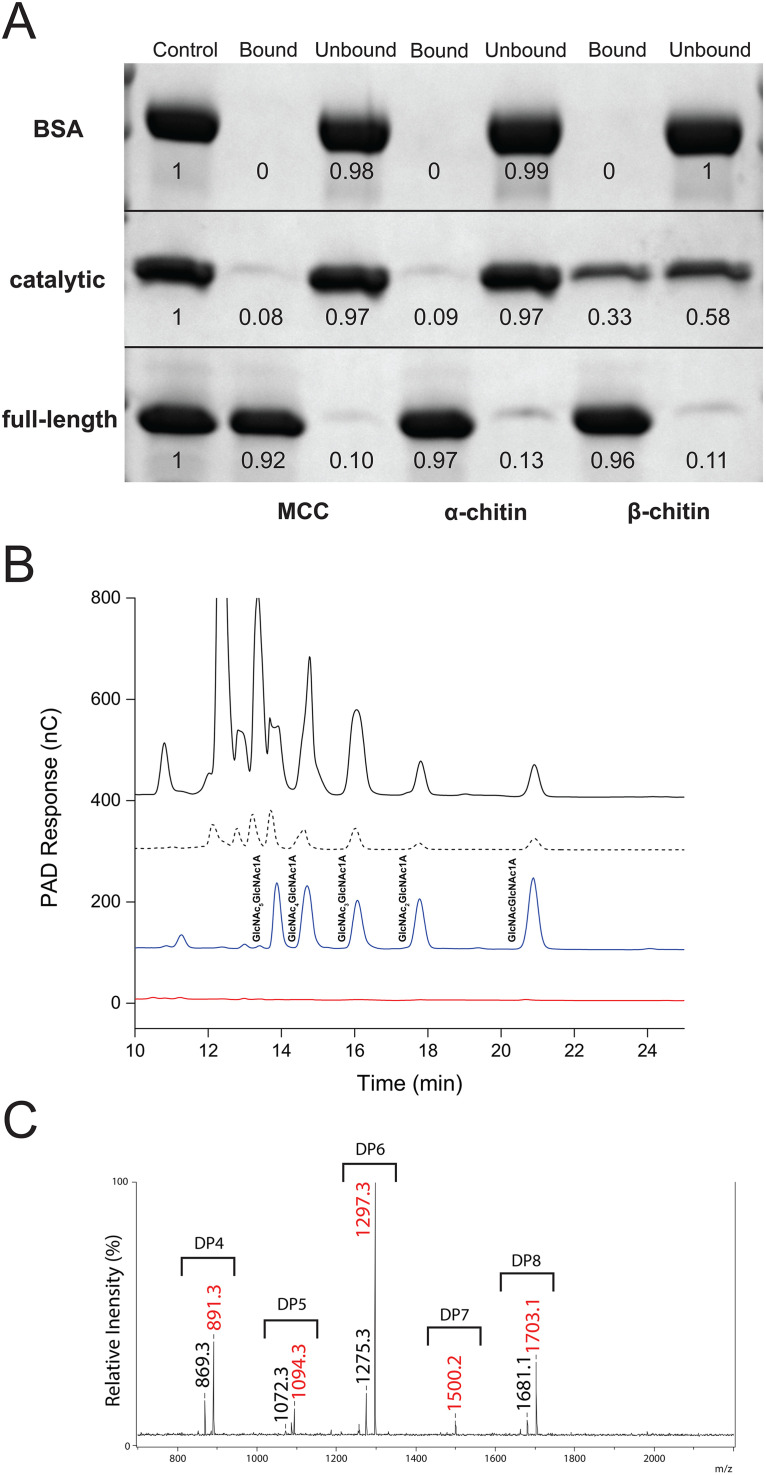
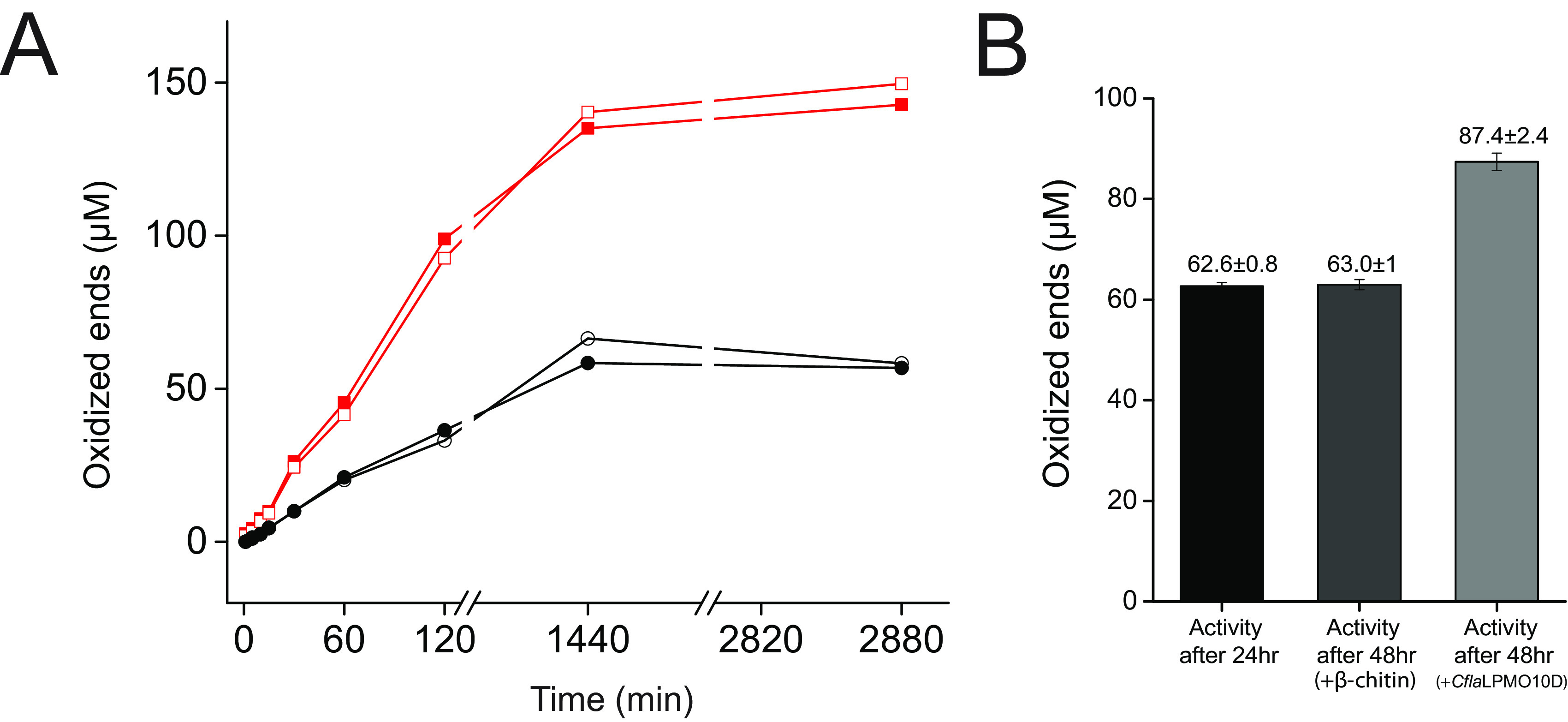
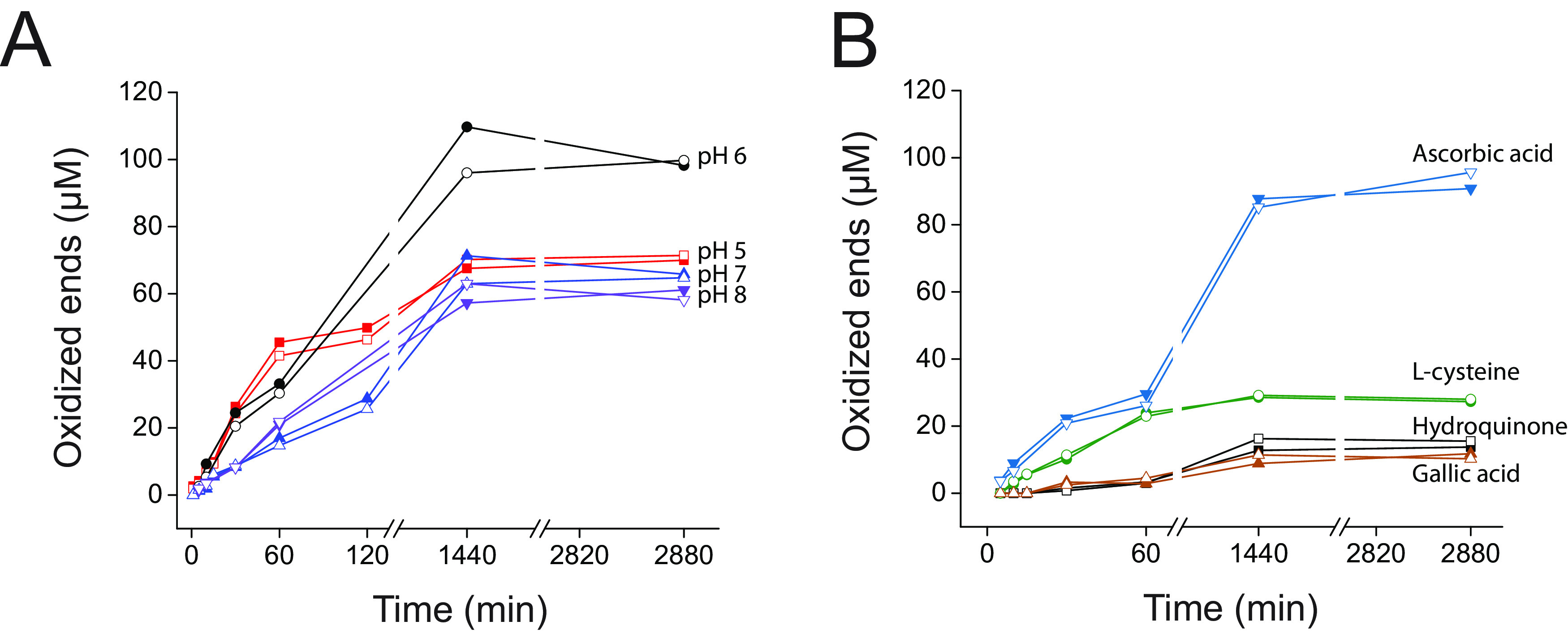
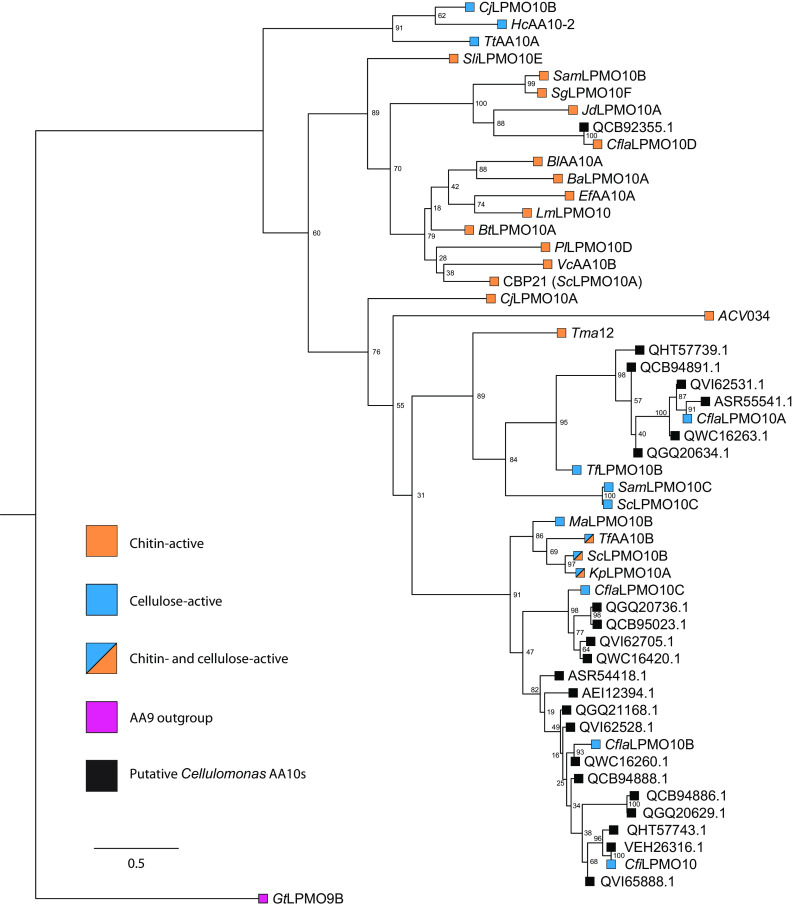
Cellulomonas flavigena is a saprotrophic bacterium that encodes, within its genome, four predicted lytic polysaccharide monooxygenases (LPMOs) from Auxiliary Activity family 10 (AA10). We showed previously that three of these cleave the plant polysaccharide cellulose by oxidation at carbon-1 (J. Li, L. Solhi, E.D. Goddard-Borger, Y. Mattieu et al., Biotechnol Biofuels 14:29, 2021, https://doi.org/10.1186/s13068-020-01860-3). Here, we present the biochemical characterization of the fourth C. flavigena AA10 member (CflaLPMO10D) as a chitin-active LPMO. Both the full-length CflaLPMO10D-Carbohydrate-Binding Module family 2 (CBM2) and catalytic module-only proteins were produced in Escherichia coli using the native general secretory (Sec) signal peptide. To quantify chitinolytic activity, we developed a high-performance anion-exchange chromatography-pulsed amperometric detection (HPAEC-PAD) method as an alternative to the established hydrophilic interaction liquid ion chromatography coupled with UV detection (HILIC-UV) method for separation and detection of released oxidized chito-oligosaccharides. Using this method, we demonstrated that CflaLPMO10D is strictly active on the β-allomorph of chitin, with optimal activity at pH 5 to 6 and a preference for ascorbic acid as the reducing agent. We also demonstrated the importance of the CBM2 member for both mediating enzyme localization to substrates and prolonging LPMO activity. Together with previous work, the present study defines the distinct substrate specificities of the suite of C. flavigena AA10 members. Notably, a cross-genome survey of AA10 members indicated that chitinolytic LPMOs are, in fact, rare among Cellulomonas bacteria. IMPORTANCE Species from the genus Cellulomonas have a long history of study due to their roles in biomass recycling in nature and corresponding potential as sources of enzymes for biotechnological applications. Although Cellulomonas species are more commonly associated with the cleavage and utilization of plant cell wall polysaccharides, here, we show that C. flavigena produces a unique lytic polysaccharide monooxygenase with activity on β-chitin, which is found, for example, in arthropods. The limited distribution of orthologous chitinolytic LPMOs suggests adaptation of individual cellulomonads to specific nutrient niches present in soil ecosystems. This research provides new insight into the biochemical specificity of LPMOs in Cellulomonas species and related bacteria, and it raises new questions about the physiological function of these enzymes.
Keywords: AA10; CBM2; Cellulomonas flavigena; HPAEC-PAD; LPMO; carbohydrate-binding module; chitin; lytic polysaccharide monooxygenase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Fernando LD, Dickwella Widanage MC, Penfield J, Lipton AS, Washton N, Latgé J-P, Wang P, Zhang L, Wang T. 2021. Structural polymorphism of chitin and chitosan in fungal cell walls from solid-state NMR and principal component analysis. Front Mol Biosci 8:727053. 10.3389/fmolb.2021.727053. - DOI - PMC - PubMed
-
- Peniche C, Argüelles-Monal W, Goycoolea FM. 2008. Chitin and chitosan: major sources, properties and applications, p 517–542. In Belgacem MN, Gandini A (ed), Monomers, polymers and composites from renewable resources. Elsevier, Oxford, UK.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
