

A Multitrait Locus Regulates Sarbecovirus Pathogenesis
- PMID: 35862771
- PMCID: PMC9426612
- DOI: 10.1128/mbio.01454-22
A Multitrait Locus Regulates Sarbecovirus Pathogenesis
Abstract
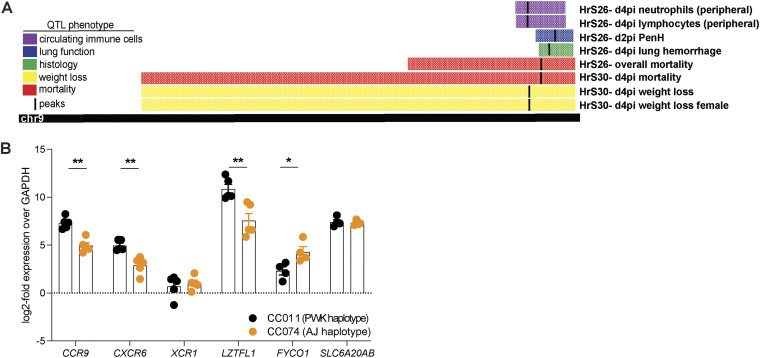
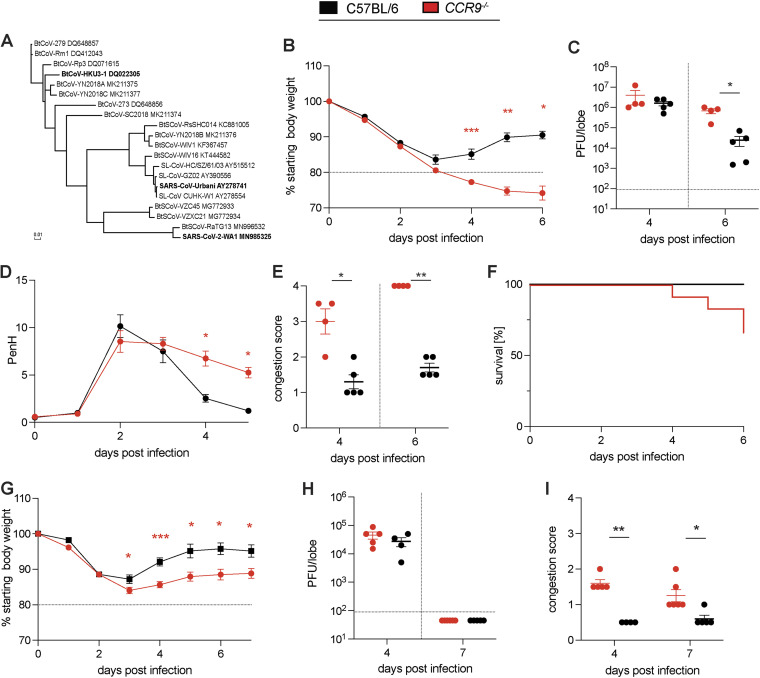
Infectious diseases have shaped the human population genetic structure, and genetic variation influences the susceptibility to many viral diseases. However, a variety of challenges have made the implementation of traditional human Genome-wide Association Studies (GWAS) approaches to study these infectious outcomes challenging. In contrast, mouse models of infectious diseases provide an experimental control and precision, which facilitates analyses and mechanistic studies of the role of genetic variation on infection. Here we use a genetic mapping cross between two distinct Collaborative Cross mouse strains with respect to severe acute respiratory syndrome coronavirus (SARS-CoV) disease outcomes. We find several loci control differential disease outcome for a variety of traits in the context of SARS-CoV infection. Importantly, we identify a locus on mouse chromosome 9 that shows conserved synteny with a human GWAS locus for SARS-CoV-2 severe disease. We follow-up and confirm a role for this locus, and identify two candidate genes, CCR9 and CXCR6, that both play a key role in regulating the severity of SARS-CoV, SARS-CoV-2, and a distantly related bat sarbecovirus disease outcomes. As such we provide a template for using experimental mouse crosses to identify and characterize multitrait loci that regulate pathogenic infectious outcomes across species. IMPORTANCE Host genetic variation is an important determinant that predicts disease outcomes following infection. In the setting of highly pathogenic coronavirus infections genetic determinants underlying host susceptibility and mortality remain unclear. To elucidate the role of host genetic variation on sarbecovirus pathogenesis and disease outcomes, we utilized the Collaborative Cross (CC) mouse genetic reference population as a model to identify susceptibility alleles to SARS-CoV and SARS-CoV-2 infections. Our findings reveal that a multitrait loci found in chromosome 9 is an important regulator of sarbecovirus pathogenesis in mice. Within this locus, we identified and validated CCR9 and CXCR6 as important regulators of host disease outcomes. Specifically, both CCR9 and CXCR6 are protective against severe SARS-CoV, SARS-CoV-2, and SARS-related HKU3 virus disease in mice. This chromosome 9 multitrait locus may be important to help identify genes that regulate coronavirus disease outcomes in humans.
Keywords: SARS-CoV-2; collaborative cross; host response; pathogenesis; sarbecoviruses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
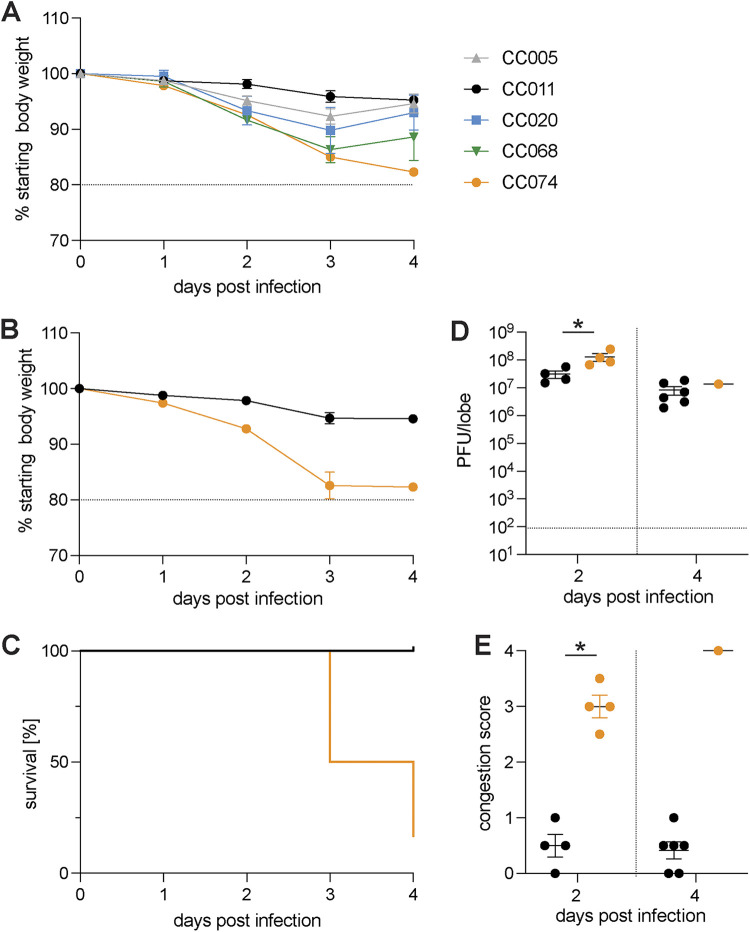
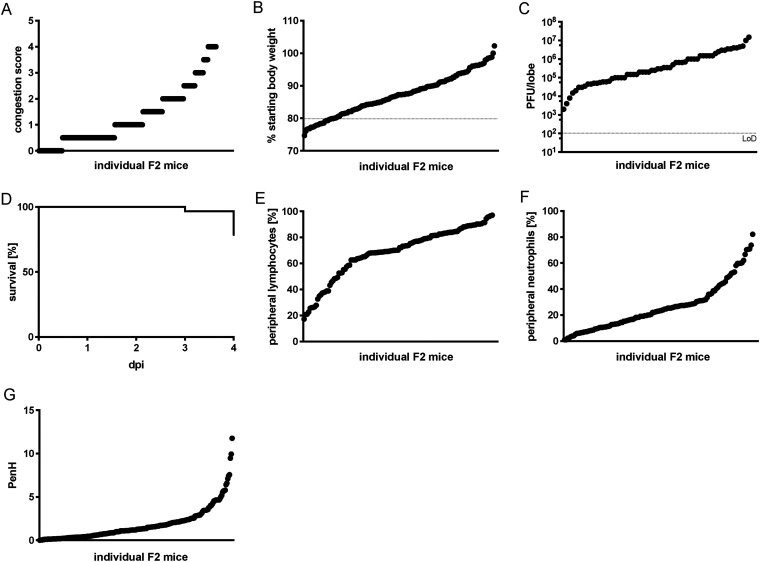
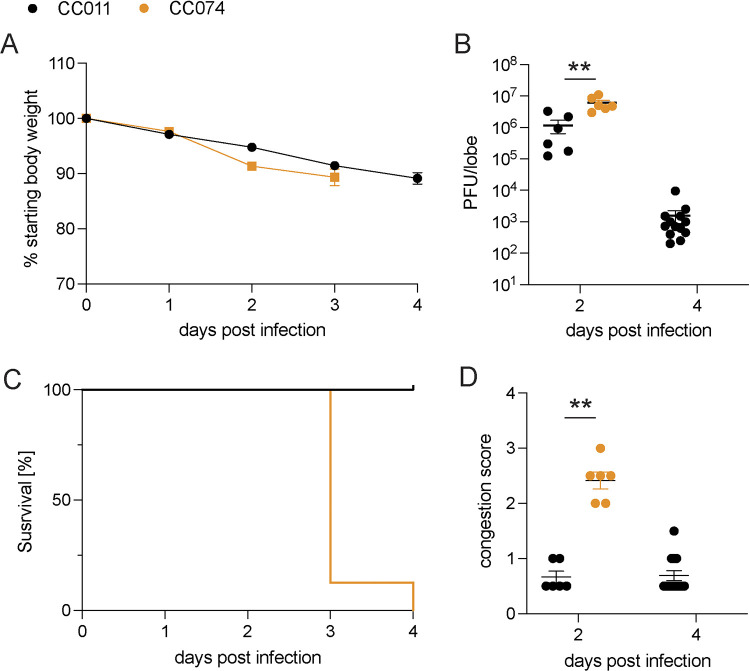
Update of
-
A Multitrait Locus Regulates Sarbecovirus Pathogenesis.bioRxiv [Preprint]. 2022 Jun 2:2022.06.01.494461. doi: 10.1101/2022.06.01.494461. bioRxiv. 2022. Update in: mBio. 2022 Aug 30;13(4):e0145422. doi: 10.1128/mbio.01454-22. PMID: 35677067 Free PMC article. Updated. Preprint.
References
-
- McLaren PJ, Coulonges C, Bartha I, Lenz TL, Deutsch AJ, Bashirova A, Buchbinder S, Carrington MN, Cossarizza A, Dalmau J, De Luca A, Goedert JJ, Gurdasani D, Haas DW, Herbeck JT, Johnson EO, Kirk GD, Lambotte O, Luo M, Mallal S, van Manen D, Martinez-Picado J, Meyer L, Miro JM, Mullins JI, Obel N, Poli G, Sandhu MS, Schuitemaker H, Shea PR, Theodorou I, Walker BD, Weintrob AC, Winkler CA, Wolinsky SM, Raychaudhuri S, Goldstein DB, Telenti A, de Bakker PIW, Zagury J-F, Fellay J. 2015. Polymorphisms of large effect explain the majority of the host genetic contribution to variation of HIV-1 virus load. Proc Natl Acad Sci USA 112:14658–14663. doi: 10.1073/pnas.1514867112. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
