

Evaluation of 3-Deoxy-D-Arabino-Heptulosonate 7-Phosphate Synthase (DAHPS) as a Vulnerable Target in Mycobacterium tuberculosis
- PMID: 35862980
- PMCID: PMC9430761
- DOI: 10.1128/spectrum.00728-22
Evaluation of 3-Deoxy-D-Arabino-Heptulosonate 7-Phosphate Synthase (DAHPS) as a Vulnerable Target in Mycobacterium tuberculosis
Abstract
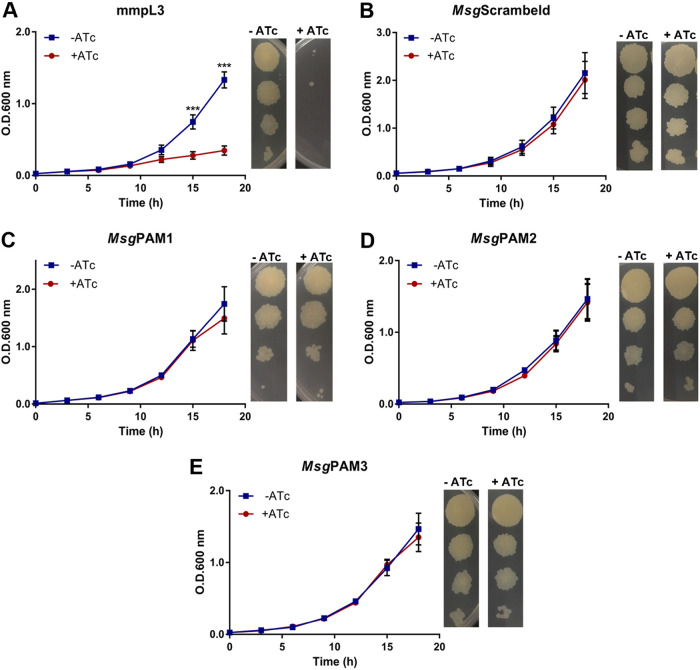
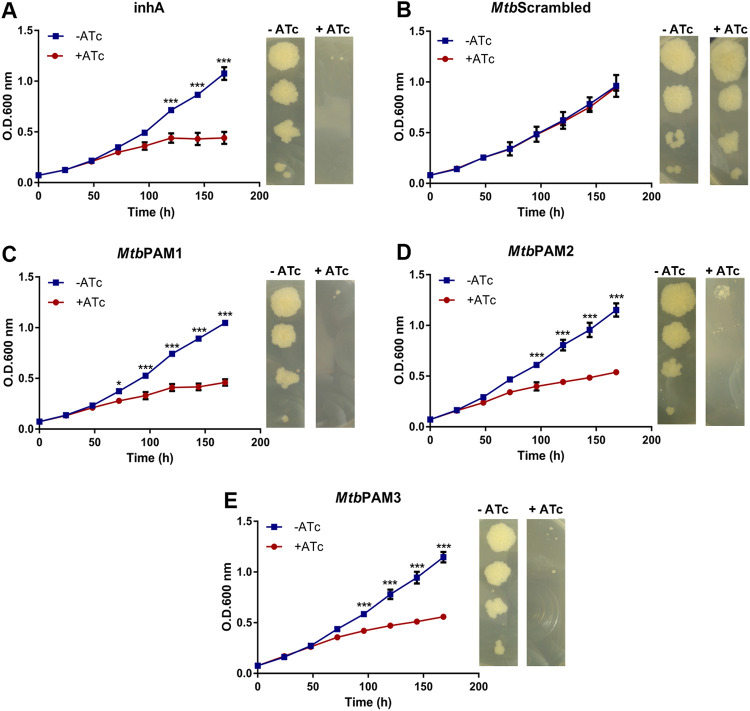
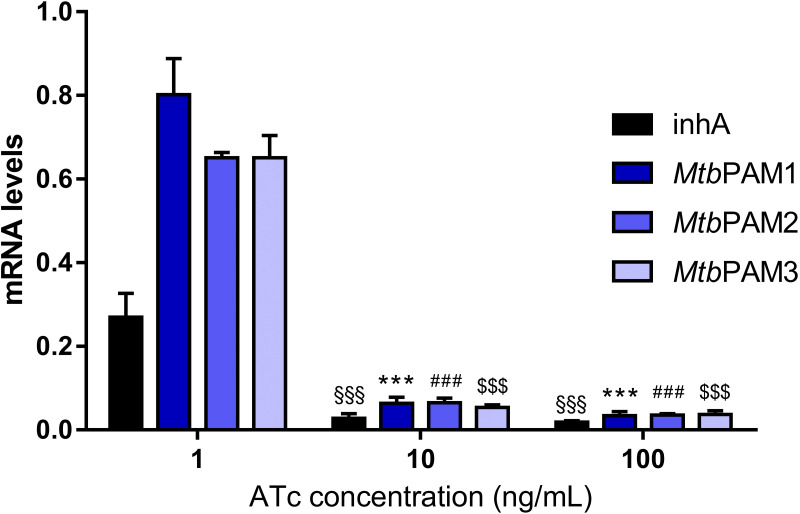
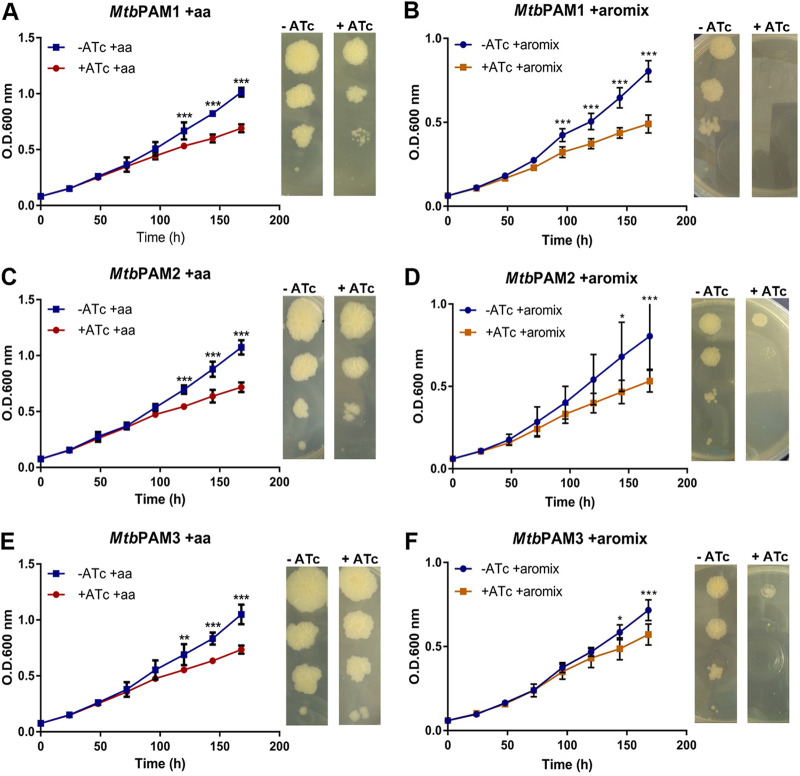
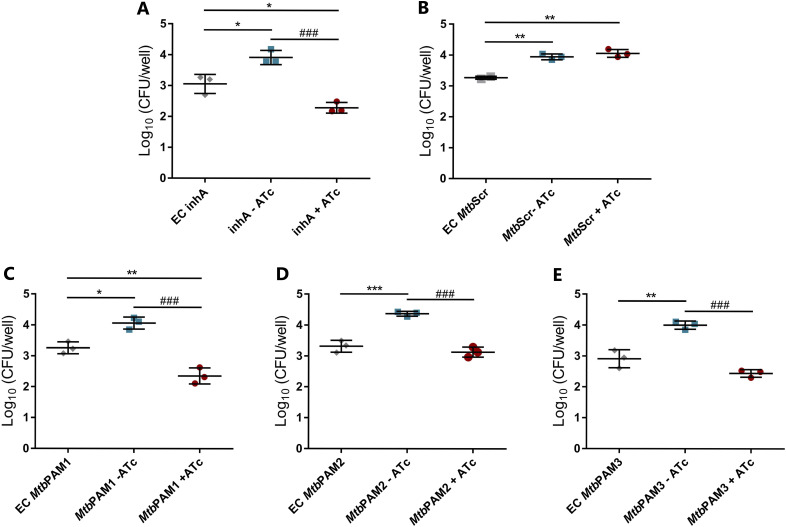
Tuberculosis (TB) remains one of the leading causes of death due to a single pathogen. The emergence and proliferation of multidrug-resistant (MDR-TB) and extensively drug-resistant strains (XDR-TB) represent compelling reasons to invest in the pursuit of new anti-TB agents. The shikimate pathway, responsible for chorismate biosynthesis, which is a precursor of important aromatic compounds, is required for Mycobacterium tuberculosis growth. The enzyme 3-deoxy-d-arabino-heptulosonate 7-phosphate synthase (MtbDAHPS) catalyzes the first step in the shikimate pathway and it is an attractive target for anti-tubercular agents. Here, we used a CRISPRi system to evaluate the DAHPS as a vulnerable target in M. tuberculosis. The silencing of aroG significantly reduces the M. tuberculosis growth in both rich medium and, especially, in infected murine macrophages. The supplementation with amino acids was only able to partially rescue the growth of bacilli, whereas the Aro supplement (aromix) was enough to sustain the bacterial growth at lower rates. This study shows that MtbDAHPS protein is vulnerable and, therefore, an attractive target to develop new anti-TB agents. In addition, the study contributes to a better understanding of the biosynthesis of aromatic compounds and the bacillus physiology. IMPORTANCE Determining the vulnerability of a potential target allows us to assess whether its partial inhibition will impact bacterial growth. Here, we evaluated the vulnerability of the enzyme 3-deoxy-d-arabino-heptulosonate 7-phosphate synthase (DAHPS) from M. tuberculosis by silencing the DAHPS-coding aroG gene in different contexts. These results could lead to the development of novel and potent anti-tubercular agents in the near future.
Keywords: CRISPRi; aroG; macrophages; shikimate pathway; vulnerability.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- WHO. 2021. Global tuberculosis report 2021. World Health Organization.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
