

The long road of functional recruitment-The evolution of a gene duplicate to pyrrolizidine alkaloid biosynthesis in the morning glories (Convolvulaceae)
- PMID: 35865076
- PMCID: PMC9295680
- DOI: 10.1002/pld3.420
The long road of functional recruitment-The evolution of a gene duplicate to pyrrolizidine alkaloid biosynthesis in the morning glories (Convolvulaceae)
Abstract
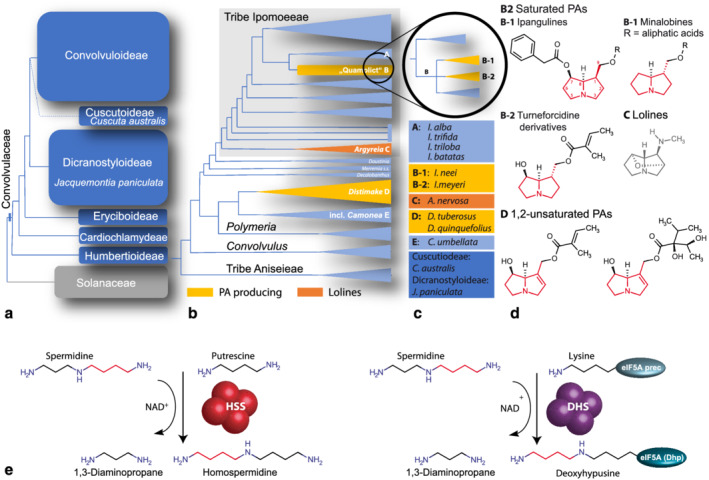
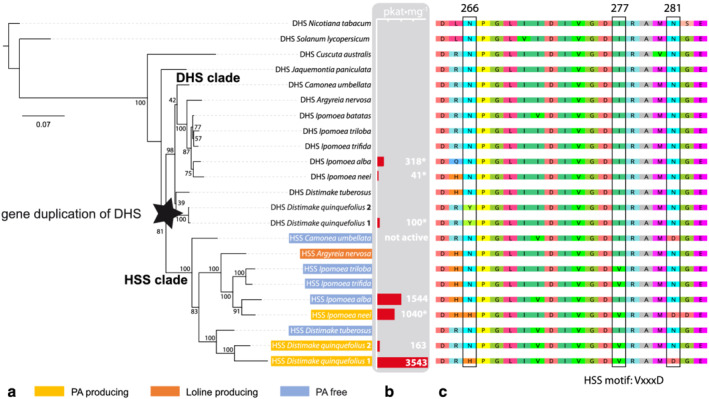
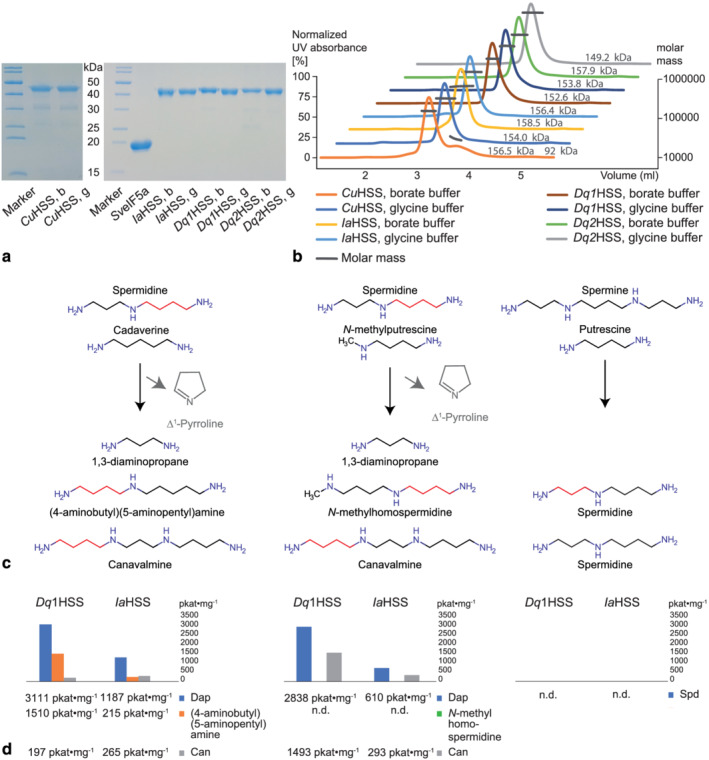
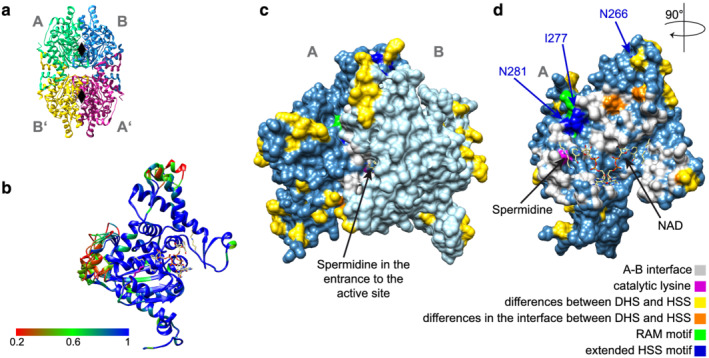
In plants, homospermidine synthase (HSS) is a pathway-specific enzyme initiating the biosynthesis of pyrrolizidine alkaloids (PAs), which function as a chemical defense against herbivores. In PA-producing Convolvulaceae ("morning glories"), HSS originated from deoxyhypusine synthase at least >50 to 75 million years ago via a gene duplication event and subsequent functional diversification. To study the recruitment of this ancient gene duplicate to PA biosynthesis, the presence of putative hss gene copies in 11 Convolvulaceae species was analyzed. Additionally, various plant parts from seven of these species were screened for the presence of PAs. Although all of these species possess a putative hss copy, PAs could only be detected in roots of Ipomoea neei (Spreng.) O'Donell and Distimake quinquefolius (L.) A.R.Simões & Staples in this study. A precursor of PAs was detected in roots of Ipomoea alba L. Thus, despite sharing high sequence identities, the presence of an hss gene copy does not correlate with PA accumulation in particular species of Convolvulaceae. In vitro activity assays of the encoded enzymes revealed a broad spectrum of enzyme activity, further emphasizing a functional diversity of the hss gene copies. A recently identified HSS specific amino acid motif seems to be important for the loss of the ancestral protein function-the activation of the eukaryotic initiation factor 5A (eIF5A). Thus, the motif might be indicative for a change of function but allows not to predict the new function. This emphasizes the challenges in annotating functions for duplicates, even for duplicates from closely related species.
Keywords: Distimake; Ipomoea; deoxyhypusine synthase; gene duplication; homospermidine synthase; molecular evolution; pyrrolizidine alkaloids.
© 2022 The Authors. Plant Direct published by American Society of Plant Biologists and the Society for Experimental Biology and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Austin, D. F. (1979). An infrageneric classification for Ipomoea (Convolvulaceae). Taxon, 28(4), 359–361.
LinkOut - more resources
Full Text Sources
Miscellaneous
