

Monitoring GAPDH activity and inhibition with cysteine-reactive chemical probes
- PMID: 35866162
- PMCID: PMC9257626
- DOI: 10.1039/d2cb00091a
Monitoring GAPDH activity and inhibition with cysteine-reactive chemical probes
Abstract
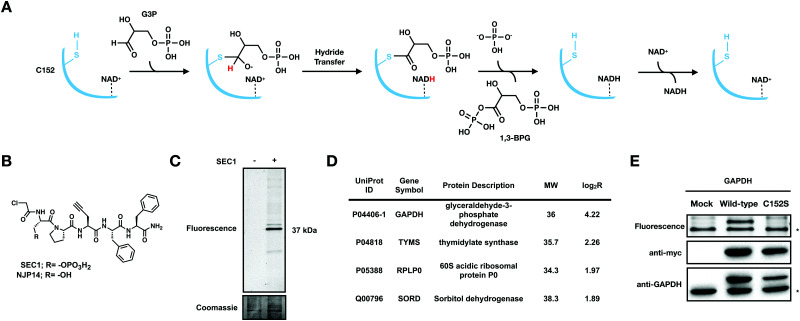
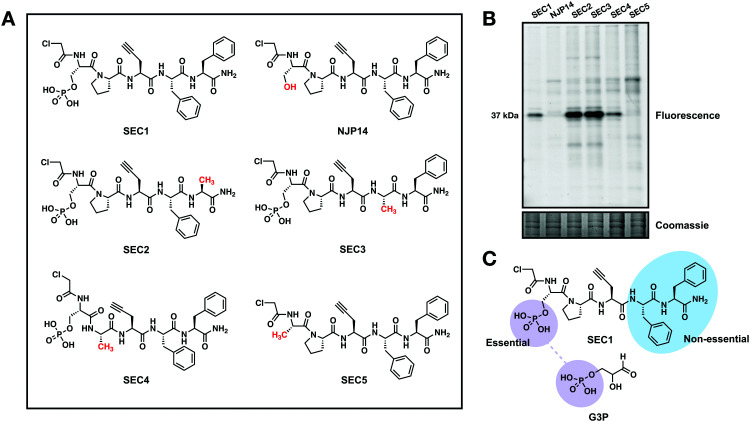
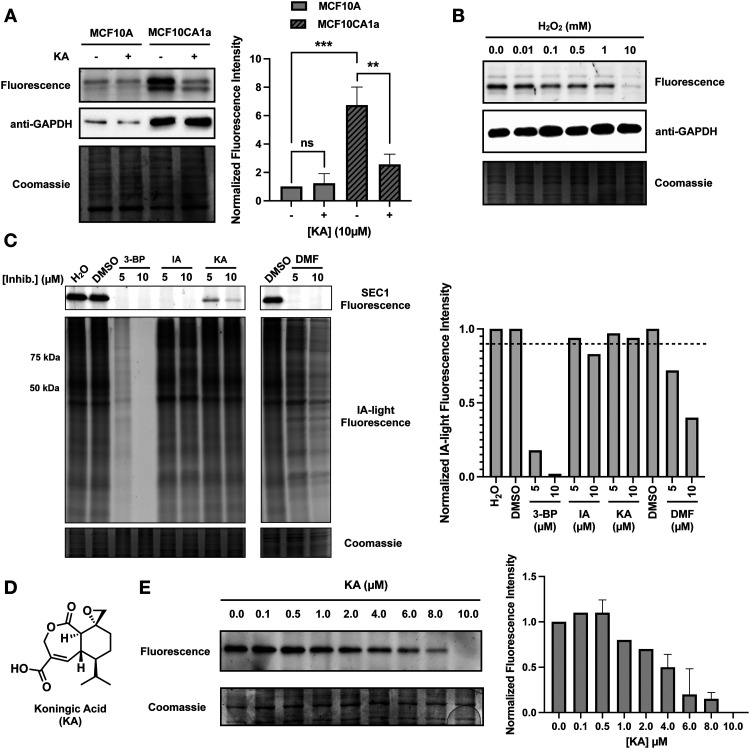
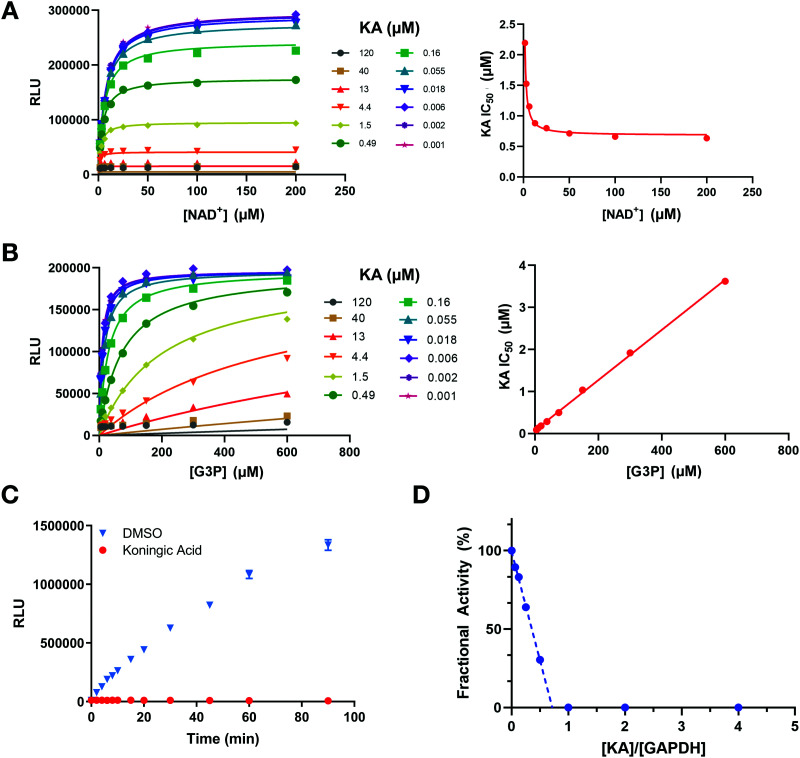
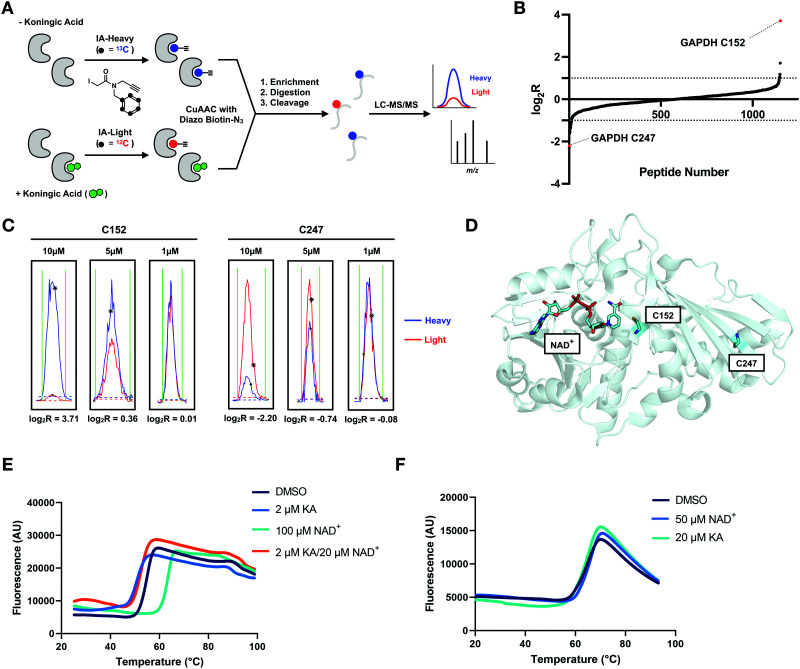
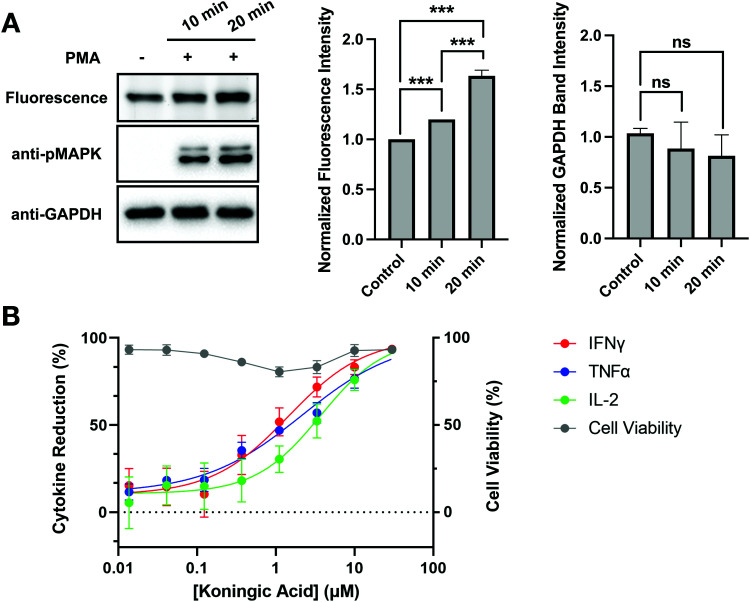
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) is a central enzyme in glycolysis that regulates the Warburg effect in cancer cells. In addition to its role in metabolism, GAPDH is also implicated in diverse cellular processes, including transcription and apoptosis. Dysregulated GAPDH activity is associated with a variety of pathologies, and GAPDH inhibitors have demonstrated therapeutic potential as anticancer and immunomodulatory agents. Given the critical role of GAPDH in pathophysiology, it is important to have access to tools that enable rapid monitoring of GAPDH activity and inhibition within a complex biological system. Here, we report an electrophilic peptide-based probe, SEC1, which covalently modifies the active-site cysteine, C152, of GAPDH to directly report on GAPDH activity within a proteome. We demonstrate the utility of SEC1 to assess changes in GAPDH activity in response to oncogenic transformation, reactive oxygen species (ROS) and small-molecule GAPDH inhibitors, including Koningic acid (KA). We then further evaluated KA, to determine the detailed mechanism of inhibition. Our mechanistic studies confirm that KA is a highly effective irreversible inhibitor of GAPDH, which acts through a NAD+-uncompetitive and G3P-competitive mechanism. Proteome-wide evaluation of the cysteine targets of KA demonstrated high selectivity for the active-site cysteine of GAPDH over other reactive cysteines within the proteome. Lastly, the therapeutic potential of KA was investigated in an autoimmune model, where treatment with KA resulted in decreased cytokine production by Th1 effector cells. Together, these studies describe methods to evaluate GAPDH activity and inhibition within a proteome, and report on the high potency and selectivity of KA as an irreversible inhibitor of GAPDH.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
The glycolytic enzyme glyceraldehyde-3-phosphate dehydrogenase works as an arsenate reductase in human red blood cells and rat liver cytosol.Toxicol Sci. 2005 Jun;85(2):859-69. doi: 10.1093/toxsci/kfi158. Epub 2005 Mar 23. Toxicol Sci. 2005. PMID: 15788719
-
The interactions of 9,10-phenanthrenequinone with glyceraldehyde-3-phosphate dehydrogenase (GAPDH), a potential site for toxic actions.Chem Biol Interact. 2005 Jun 30;155(1-2):97-110. doi: 10.1016/j.cbi.2005.05.002. Chem Biol Interact. 2005. PMID: 15950210
-
Evolved resistance to partial GAPDH inhibition results in loss of the Warburg effect and in a different state of glycolysis.J Biol Chem. 2020 Jan 3;295(1):111-124. doi: 10.1074/jbc.RA119.010903. Epub 2019 Nov 20. J Biol Chem. 2020. PMID: 31748414 Free PMC article.
-
Covalent inhibitors of GAPDH: From unspecific warheads to selective compounds.Eur J Med Chem. 2020 Dec 1;207:112740. doi: 10.1016/j.ejmech.2020.112740. Epub 2020 Aug 22. Eur J Med Chem. 2020. PMID: 32898762 Review.
-
Inhibitors of Glyceraldehyde 3-Phosphate Dehydrogenase and Unexpected Effects of Its Reduced Activity.Biochemistry (Mosc). 2019 Nov;84(11):1268-1279. doi: 10.1134/S0006297919110051. Biochemistry (Mosc). 2019. PMID: 31760917 Review.
Cited by
-
Effects of Metformin Treatment Against Endometrial Cancer Cells Cultured In Vitro or Grafted into Female Balb/C Nude Mice: Insights into Cell Response and IGF-1R and PI3K/AKT/mTOR Signaling Pathways.Cell Biochem Biophys. 2025 Jul 21. doi: 10.1007/s12013-025-01840-0. Online ahead of print. Cell Biochem Biophys. 2025. PMID: 40690138
-
Haloacetonitriles Induce Structure-Related Cellular Toxicity Through Distinct Proteome Thiol Reaction Mechanisms.ACS Environ Au. 2024 Dec 3;5(1):101-113. doi: 10.1021/acsenvironau.4c00068. eCollection 2025 Jan 15. ACS Environ Au. 2024. PMID: 39830722 Free PMC article.
-
Role of Glycolytic and Glutamine Metabolism Reprogramming on the Proliferation, Invasion, and Apoptosis Resistance through Modulation of Signaling Pathways in Glioblastoma.Int J Mol Sci. 2023 Dec 18;24(24):17633. doi: 10.3390/ijms242417633. Int J Mol Sci. 2023. PMID: 38139462 Free PMC article. Review.
References
-
- Cori C. A. Cori G. T. The carbohydrate metabolism of tumours. J. Biol. Chem. 1925;65:397–405. doi: 10.1016/S0021-9258(18)84849-9. - DOI
-
- Liberti M. V. Dai Z. Wardell S. E. Baccile J. A. Liu X. Gao X. Baldi R. Mehrmohamadi M. Johnson M. O. Madhukar N. S. Shestov A. A. Chio I. Elemento O. Rathmell J. C. Schroeder F. C. McDonnell D. P. Locasale J. W. A Predictive model for selective targeting of the Warburg effect through GAPDH inhibition with a natural product. Cell Metab. 2017;26(4):648–659. doi: 10.1016/j.cmet.2017.08.017. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials