

Principles and correction of 5'-splice site selection
- PMID: 35866748
- PMCID: PMC9311317
- DOI: 10.1080/15476286.2022.2100971
Principles and correction of 5'-splice site selection
Abstract
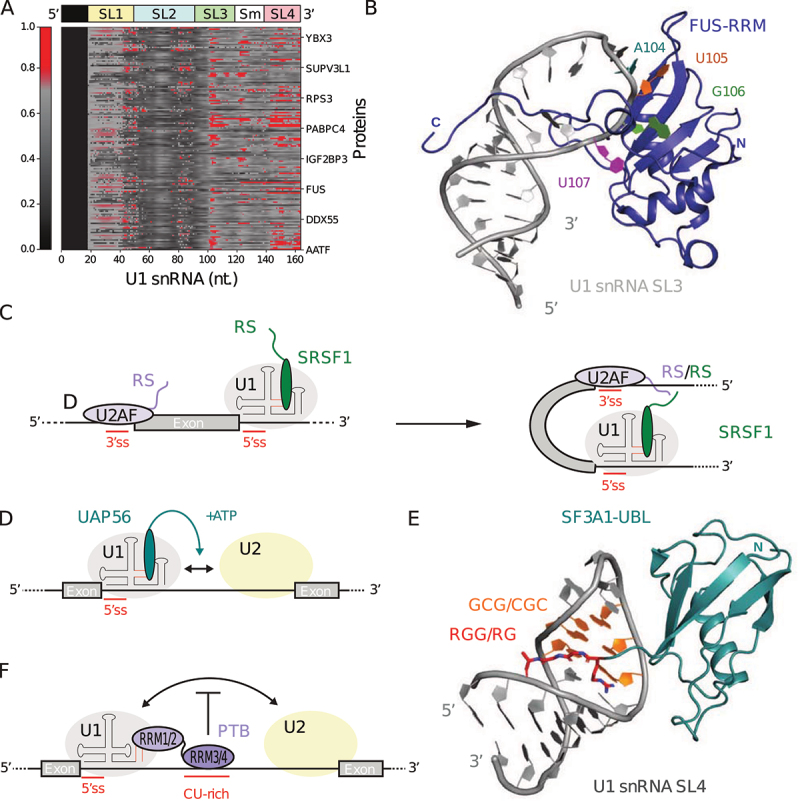
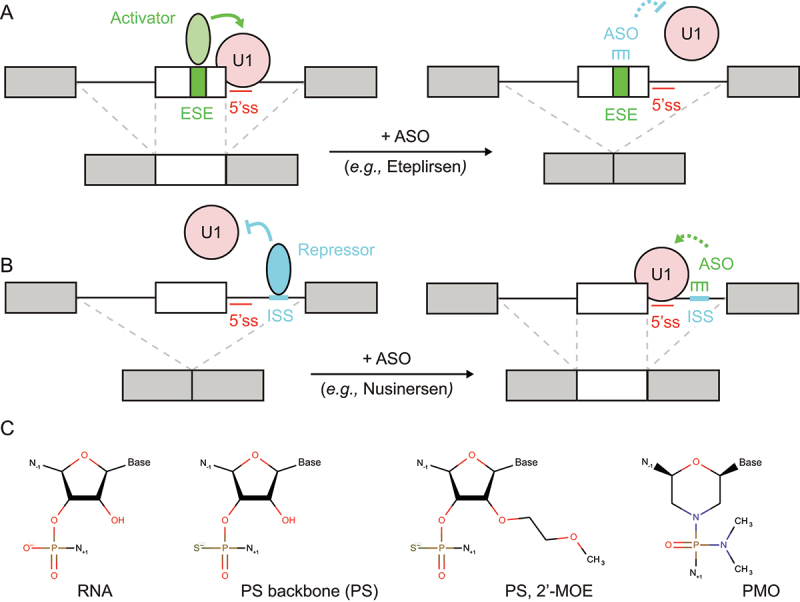
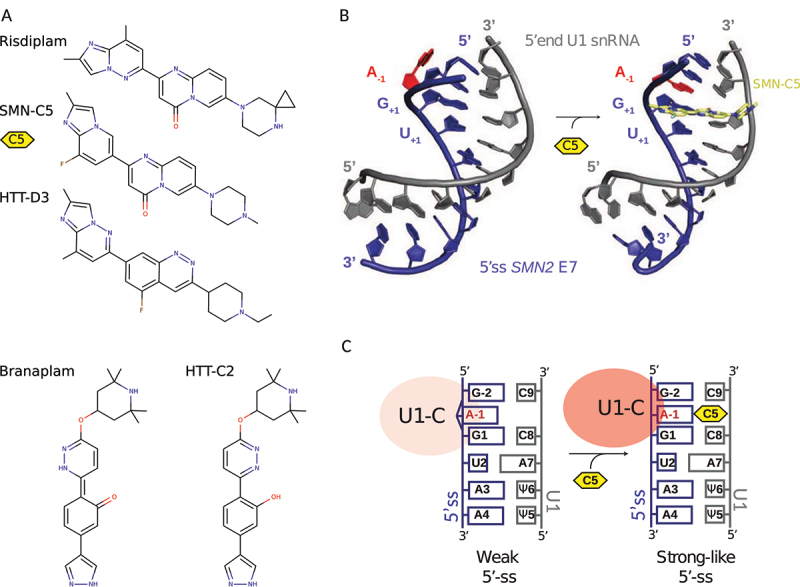
In Eukarya, immature mRNA transcripts (pre-mRNA) often contain coding sequences, or exons, interleaved by non-coding sequences, or introns. Introns are removed upon splicing, and further regulation of the retained exons leads to alternatively spliced mRNA. The splicing reaction requires the stepwise assembly of the spliceosome, a macromolecular machine composed of small nuclear ribonucleoproteins (snRNPs). This review focuses on the early stage of spliceosome assembly, when U1 snRNP defines each intron 5'-splice site (5'ss) in the pre-mRNA. We first introduce the splicing reaction and the impact of alternative splicing on gene expression regulation. Thereafter, we extensively discuss splicing descriptors that influence the 5'ss selection by U1 snRNP, such as sequence determinants, and interactions mediated by U1-specific proteins or U1 small nuclear RNA (U1 snRNA). We also include examples of diseases that affect the 5'ss selection by U1 snRNP, and discuss recent therapeutic advances that manipulate U1 snRNP 5'ss selectivity with antisense oligonucleotides and small-molecule splicing switches.
Keywords: 5’-splice site; RNA splicing; U1 snRNP; antisense oligonucleotides; splicing modifiers.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
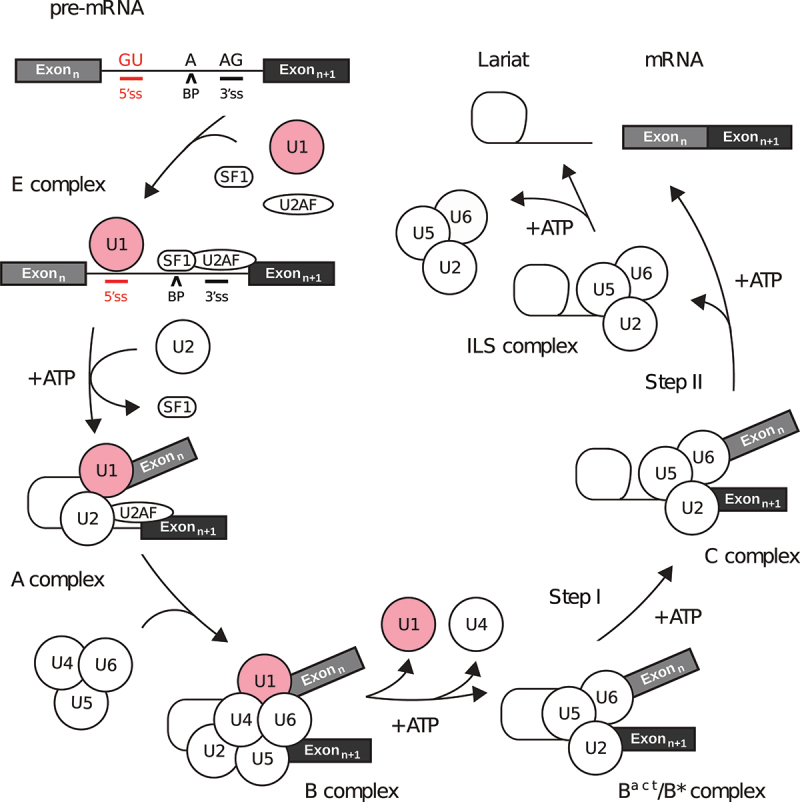
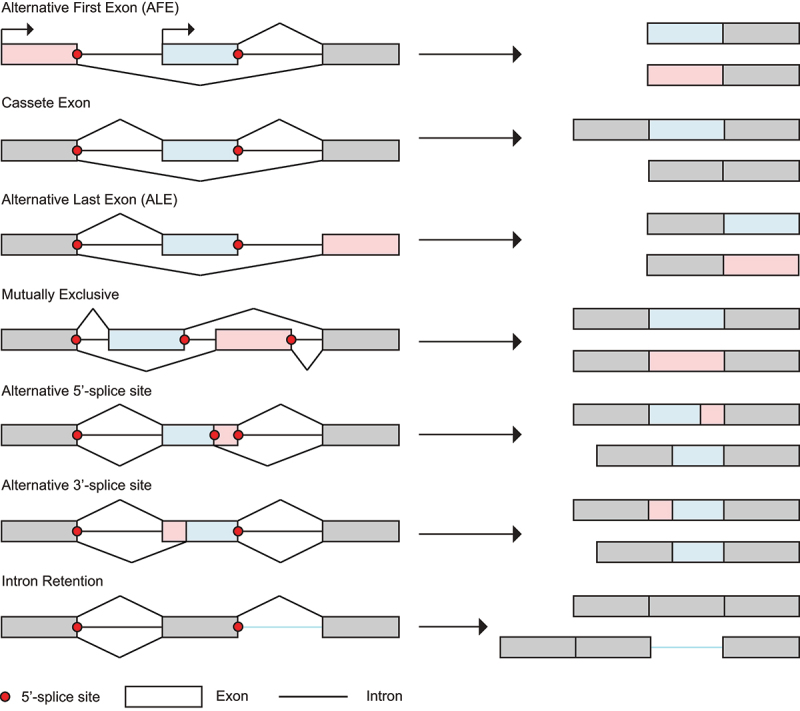
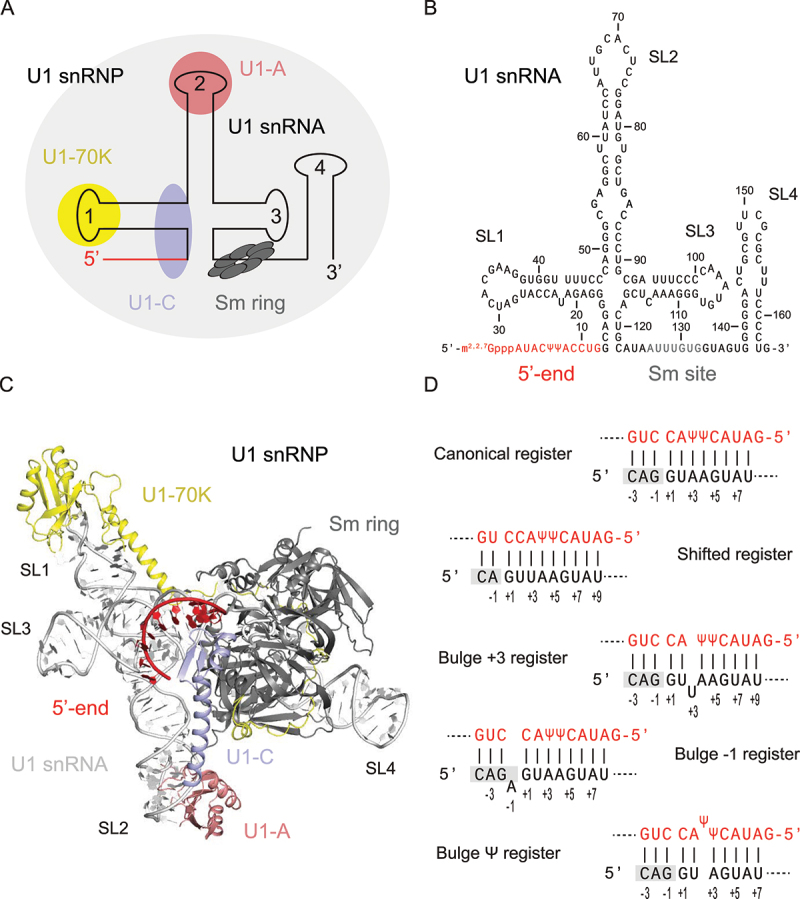
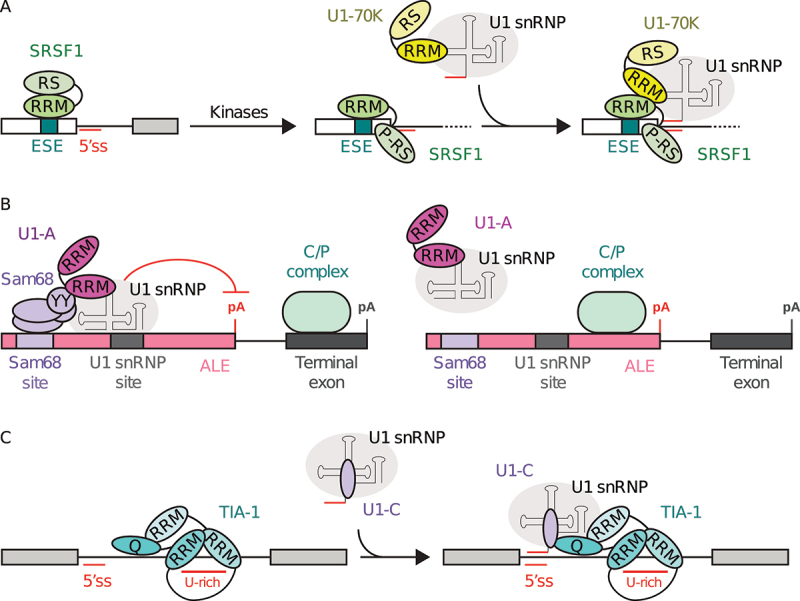
Similar articles
-
A novel role of U1 snRNP: Splice site selection from a distance.Biochim Biophys Acta Gene Regul Mech. 2019 Jun;1862(6):634-642. doi: 10.1016/j.bbagrm.2019.04.004. Epub 2019 Apr 28. Biochim Biophys Acta Gene Regul Mech. 2019. PMID: 31042550 Free PMC article. Review.
-
Selection of alternative 5' splice sites: role of U1 snRNP and models for the antagonistic effects of SF2/ASF and hnRNP A1.Mol Cell Biol. 2000 Nov;20(22):8303-18. doi: 10.1128/MCB.20.22.8303-8318.2000. Mol Cell Biol. 2000. PMID: 11046128 Free PMC article.
-
Profiling of cis- and trans-acting factors supporting noncanonical splice site activation.RNA Biol. 2021 Jan;18(1):118-130. doi: 10.1080/15476286.2020.1798111. Epub 2020 Aug 5. RNA Biol. 2021. PMID: 32693676 Free PMC article.
-
Mouse nuclear RNAi-defective 2 promotes splicing of weak 5' splice sites.RNA. 2023 Aug;29(8):1140-1165. doi: 10.1261/rna.079465.122. Epub 2023 May 3. RNA. 2023. PMID: 37137667 Free PMC article.
-
Branch site recognition by the spliceosome.RNA. 2024 Oct 16;30(11):1397-1407. doi: 10.1261/rna.080198.124. RNA. 2024. PMID: 39187383 Free PMC article. Review.
Cited by
-
Comparative Proteomic Analysis Provides New Insights into the Molecular Basis of Thermal-Induced Parthenogenesis in Silkworm (Bombyx mori).Insects. 2023 Jan 28;14(2):134. doi: 10.3390/insects14020134. Insects. 2023. PMID: 36835703 Free PMC article.
-
A sequential binding mechanism for 5' splice site recognition and modulation for the human U1 snRNP.Nat Commun. 2024 Oct 10;15(1):8776. doi: 10.1038/s41467-024-53124-5. Nat Commun. 2024. PMID: 39389991 Free PMC article.
-
Neuron-specific repression of alternative splicing by the conserved CELF protein UNC-75 in Caenorhabditis elegans.Genetics. 2025 Apr 17;229(4):iyaf025. doi: 10.1093/genetics/iyaf025. Genetics. 2025. PMID: 40059624 Free PMC article.
-
A Sequential Binding Mechanism for 5' Splice Site Recognition and Modulation for the Human U1 snRNP.bioRxiv [Preprint]. 2024 Apr 18:2024.04.18.590139. doi: 10.1101/2024.04.18.590139. bioRxiv. 2024. Update in: Nat Commun. 2024 Oct 10;15(1):8776. doi: 10.1038/s41467-024-53124-5. PMID: 38659798 Free PMC article. Updated. Preprint.
-
RNA therapeutics.RNA Biol. 2024 Jan;21(1):1-2. doi: 10.1080/15476286.2022.2161704. Epub 2023 Jan 11. RNA Biol. 2024. PMID: 36629444 Free PMC article. No abstract available.
References
-
- Alberts B, Johnson A, and Lewis J.. Molecular biology of the cell. 4th ed. New York: Garland Science. 2003.
-
- Hombach S, Kretz M. Non-coding RNAs: classification, biology and functioning. Adv Exp Med Biol. 2016;937:3–17. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources