

Motor guidance by long-range communication on the microtubule highway
- PMID: 35867749
- PMCID: PMC9282251
- DOI: 10.1073/pnas.2120193119
Motor guidance by long-range communication on the microtubule highway
Abstract
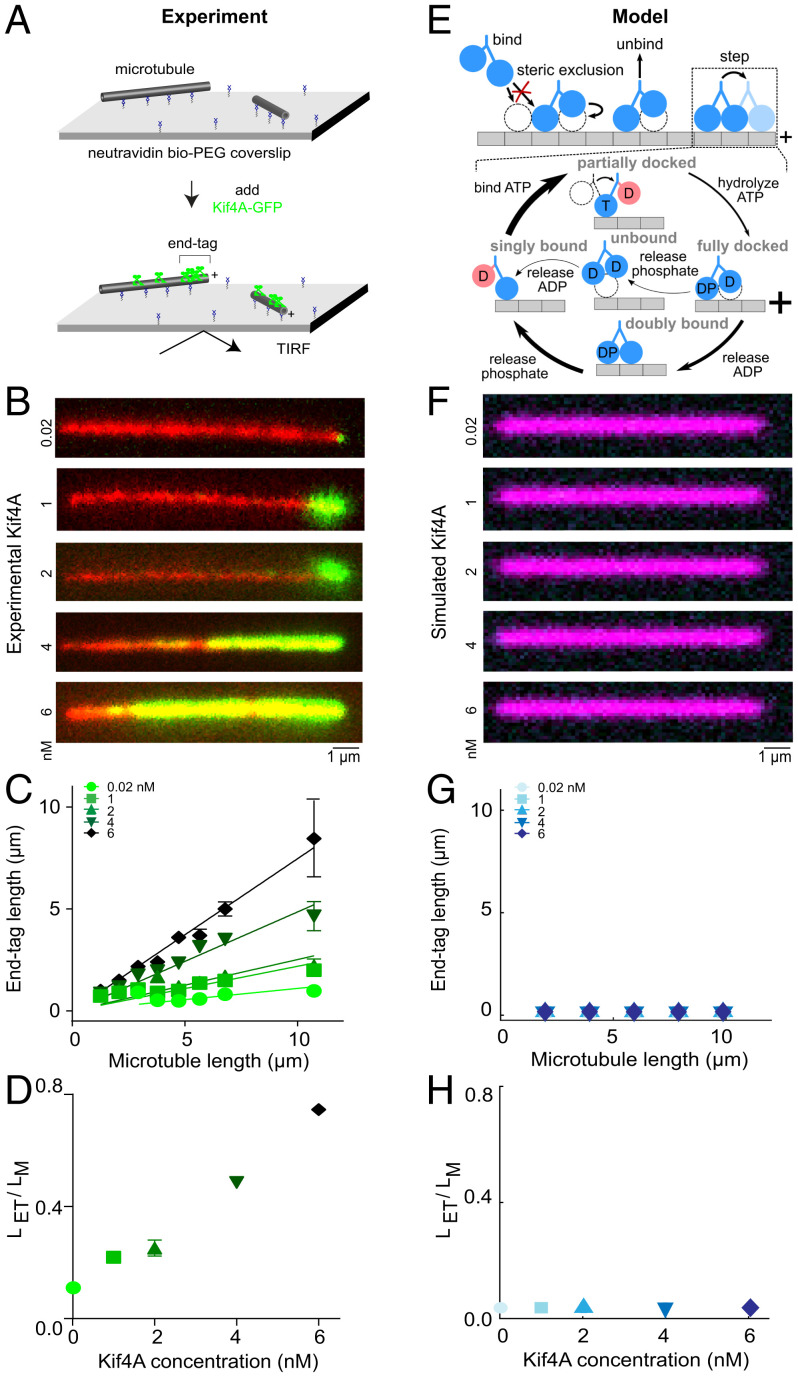
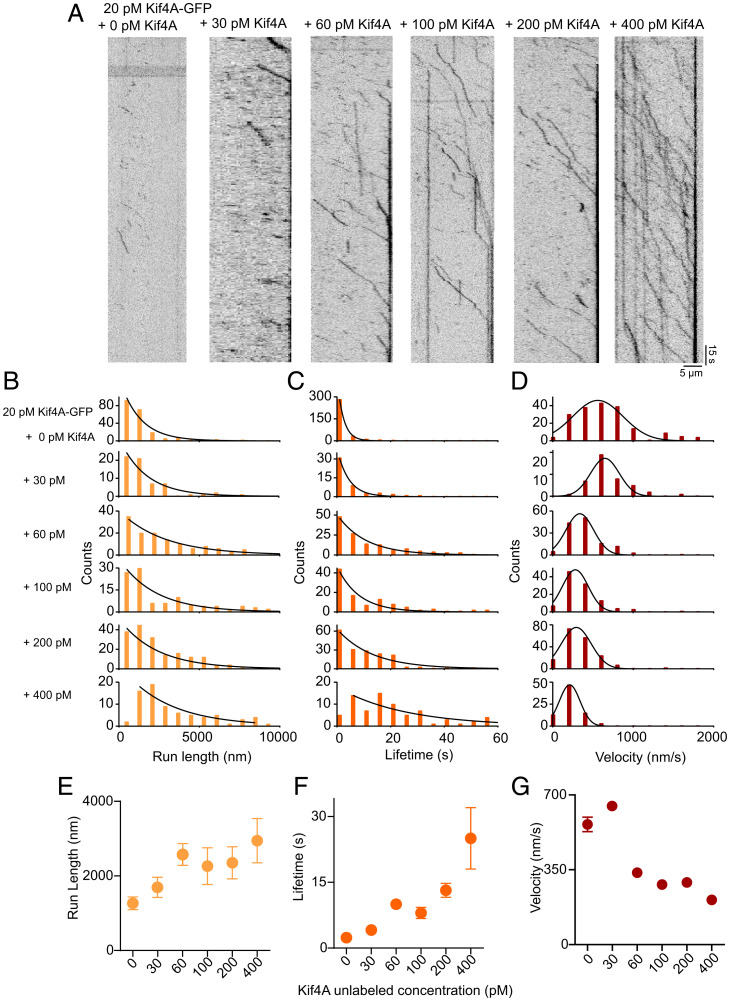
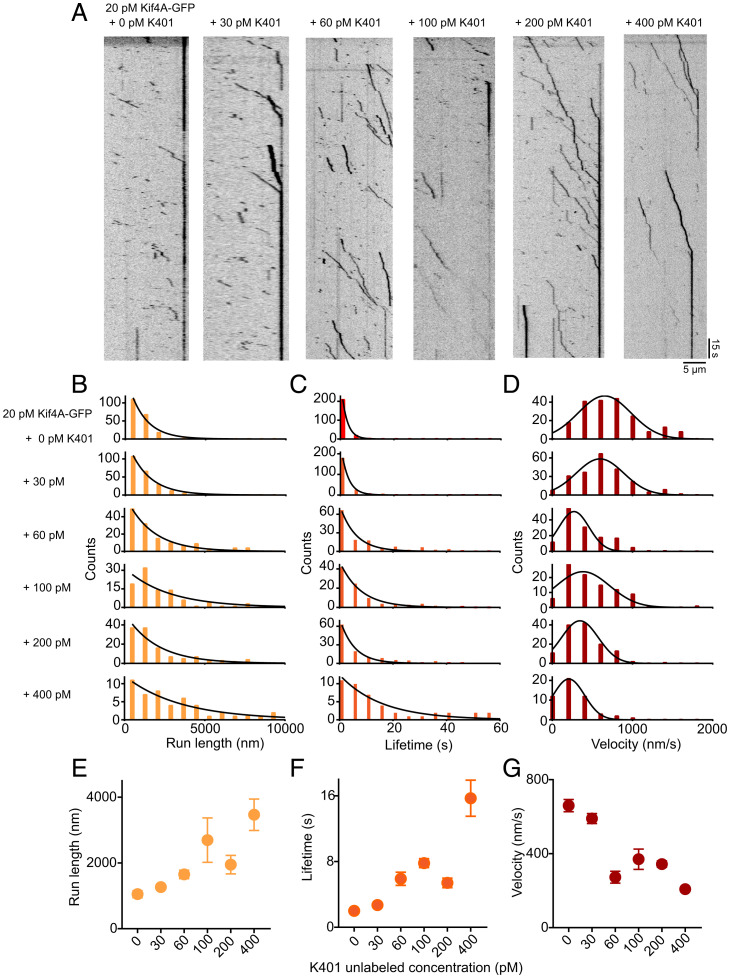
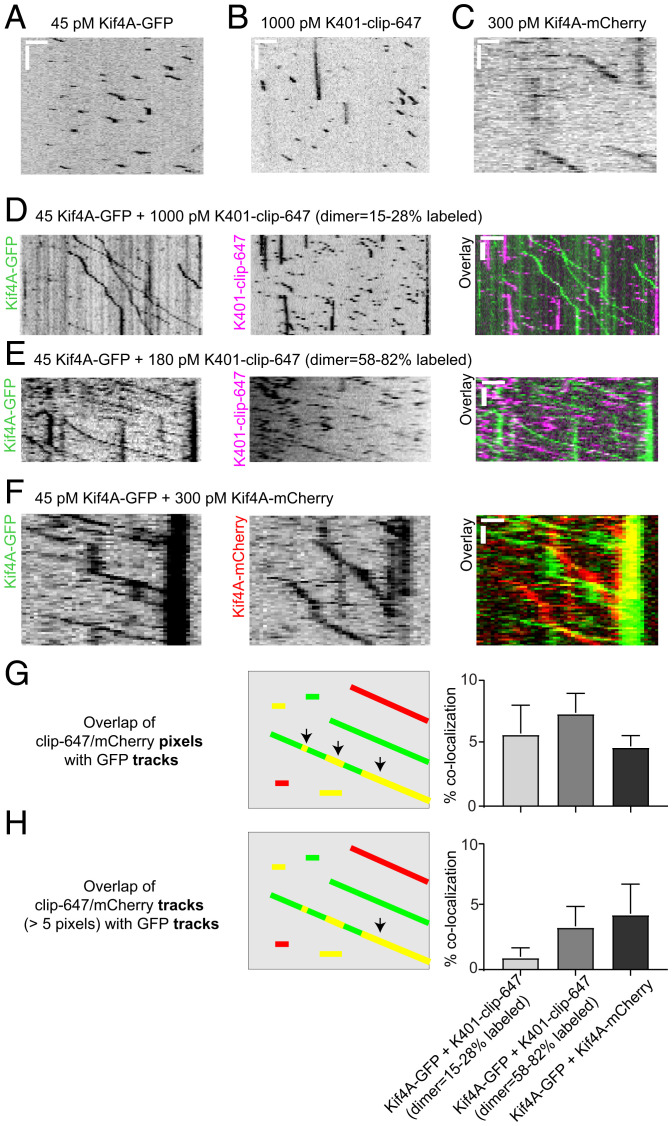
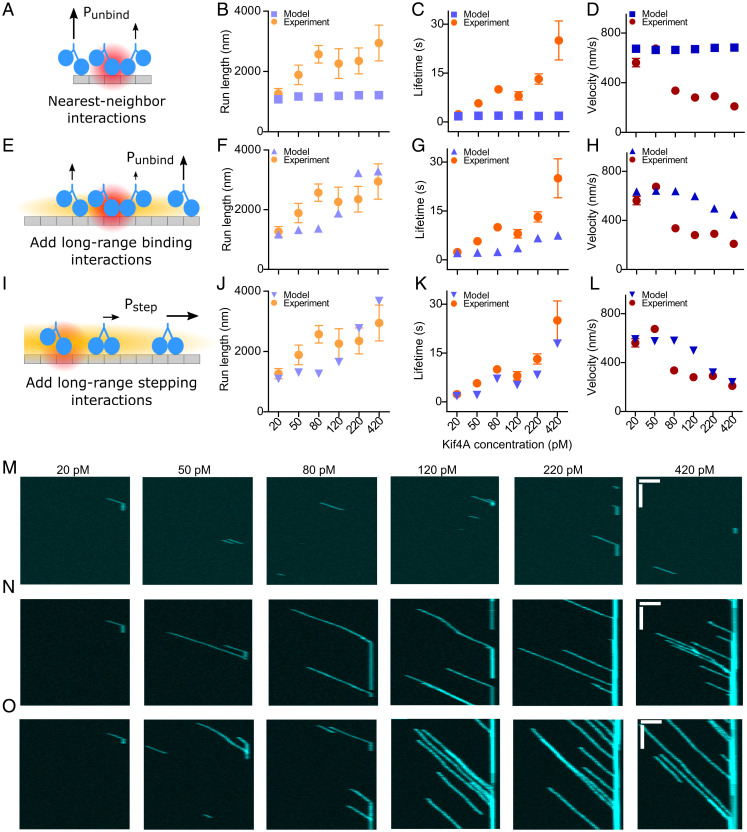
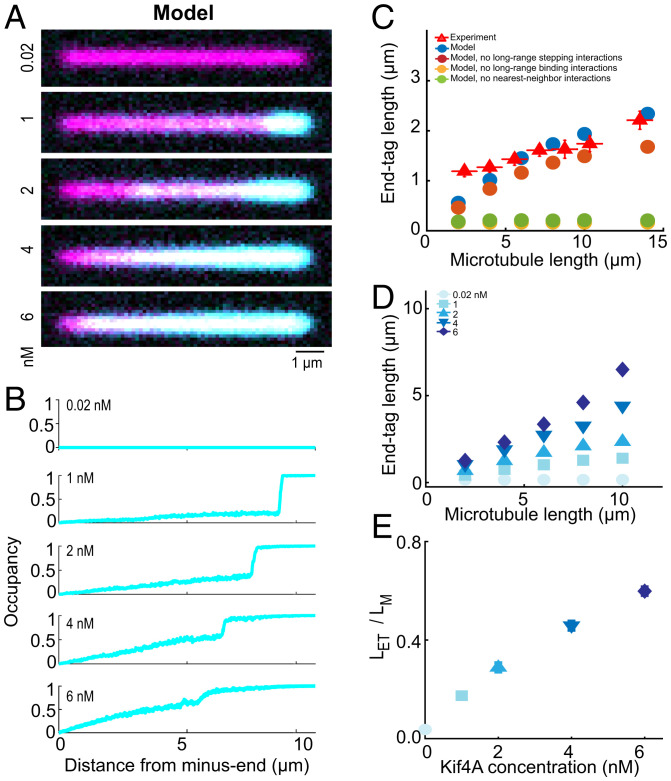
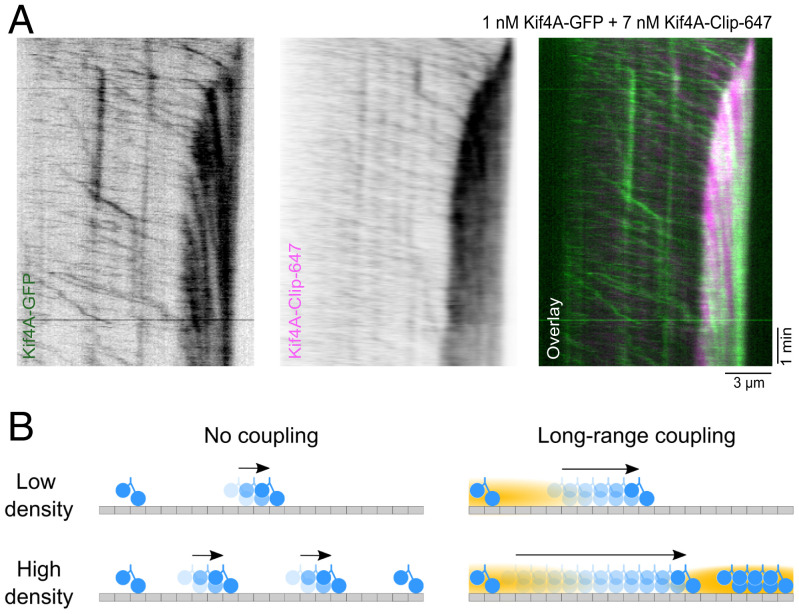
Coupling of motor proteins within arrays drives muscle contraction, flagellar beating, chromosome segregation, and other biological processes. Current models of motor coupling invoke either direct mechanical linkage or protein crowding, which rely on short-range motor-motor interactions. In contrast, coupling mechanisms that act at longer length scales remain largely unexplored. Here we report that microtubules can physically couple motor movement in the absence of detectable short-range interactions. The human kinesin-4 Kif4A changes the run length and velocity of other motors on the same microtubule in the dilute binding limit, when approximately 10-nm-sized motors are much farther apart than the motor size. This effect does not depend on specific motor-motor interactions because similar changes in Kif4A motility are induced by kinesin-1 motors. A micrometer-scale attractive interaction potential between motors is sufficient to recreate the experimental results in a biophysical model. Unexpectedly, our theory suggests that long-range microtubule-mediated coupling affects not only binding kinetics but also motor mechanochemistry. Therefore, the model predicts that motors can sense and respond to motors bound several micrometers away on a microtubule. Our results are consistent with a paradigm in which long-range motor interactions along the microtubule enable additional forms of collective motor behavior, possibly due to changes in the microtubule lattice.
Keywords: cytoskeleton; kinesin; microtubules; motors.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Dynamic kinesin-1 clustering on microtubules due to mutually attractive interactions.Phys Biol. 2008 Nov 24;5(4):046004. doi: 10.1088/1478-3975/5/4/046004. Phys Biol. 2008. PMID: 19029597
-
Kinesin-14 motors participate in a force balance at microtubule plus-ends to regulate dynamic instability.Proc Natl Acad Sci U S A. 2022 Feb 22;119(8):e2108046119. doi: 10.1073/pnas.2108046119. Proc Natl Acad Sci U S A. 2022. PMID: 35173049 Free PMC article.
-
Crowding and Pausing Strongly Affect Dynamics of Kinesin-1 Motors along Microtubules.Biophys J. 2018 Sep 18;115(6):1068-1081. doi: 10.1016/j.bpj.2018.07.017. Epub 2018 Jul 25. Biophys J. 2018. PMID: 30146266 Free PMC article.
-
Kinesin-8 motors: regulation of microtubule dynamics and chromosome movements.Chromosoma. 2020 Jun;129(2):99-110. doi: 10.1007/s00412-020-00736-7. Epub 2020 May 17. Chromosoma. 2020. PMID: 32417983 Review.
-
Lucky 13-microtubule depolymerisation by kinesin-13 motors.J Cell Sci. 2006 Oct 1;119(Pt 19):3905-13. doi: 10.1242/jcs.03224. J Cell Sci. 2006. PMID: 16988025 Review.
Cited by
-
Kinesin-1-transported liposomes prefer to go straight in 3D microtubule intersections by a mechanism shared by other molecular motors.Proc Natl Acad Sci U S A. 2024 Jul 16;121(29):e2407330121. doi: 10.1073/pnas.2407330121. Epub 2024 Jul 9. Proc Natl Acad Sci U S A. 2024. PMID: 38980901 Free PMC article.
-
Mechanism and regulation of kinesin motors.Nat Rev Mol Cell Biol. 2025 Feb;26(2):86-103. doi: 10.1038/s41580-024-00780-6. Epub 2024 Oct 11. Nat Rev Mol Cell Biol. 2025. PMID: 39394463 Review.
-
Tubulin CFEOM mutations both inhibit or activate kinesin motor activity.Mol Biol Cell. 2024 Mar 1;35(3):ar32. doi: 10.1091/mbc.E23-01-0020. Epub 2024 Jan 3. Mol Biol Cell. 2024. PMID: 38170592 Free PMC article.
-
Micron-scale geometrical features of microtubules as regulators of microtubule organization.Elife. 2021 Jun 11;10:e63880. doi: 10.7554/eLife.63880. Elife. 2021. PMID: 34114950 Free PMC article. Review.
-
Microscopic interactions control a structural transition in active mixtures of microtubules and molecular motors.Proc Natl Acad Sci U S A. 2024 Jan 9;121(2):e2300174121. doi: 10.1073/pnas.2300174121. Epub 2024 Jan 4. Proc Natl Acad Sci U S A. 2024. PMID: 38175870 Free PMC article.
References
-
- Huxley H. E., Brown W., The low-angle x-ray diagram of vertebrate striated muscle and its behaviour during contraction and rigor. J. Mol. Biol. 30, 383–434 (1967). - PubMed
-
- Craig R., Woodhead J. L., Structure and function of myosin filaments. Curr. Opin. Struct. Biol. 16, 204–212 (2006). - PubMed
-
- Gibbons I. R., Rowe A. J., Dynein: A protein with adenosine triphosphatase activity from cilia. Science 149, 424–426 (1965). - PubMed
-
- Mitchison T. J., Mitchison H. M., Cell biology: How cilia beat. Nature 463, 308–309 (2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
