

Increased GABA transmission to GnRH neurons after intrahippocampal kainic acid injection in mice is sex-specific and associated with estrous cycle disruption
- PMID: 35868435
- PMCID: PMC9455811
- DOI: 10.1016/j.nbd.2022.105822
Increased GABA transmission to GnRH neurons after intrahippocampal kainic acid injection in mice is sex-specific and associated with estrous cycle disruption
Abstract
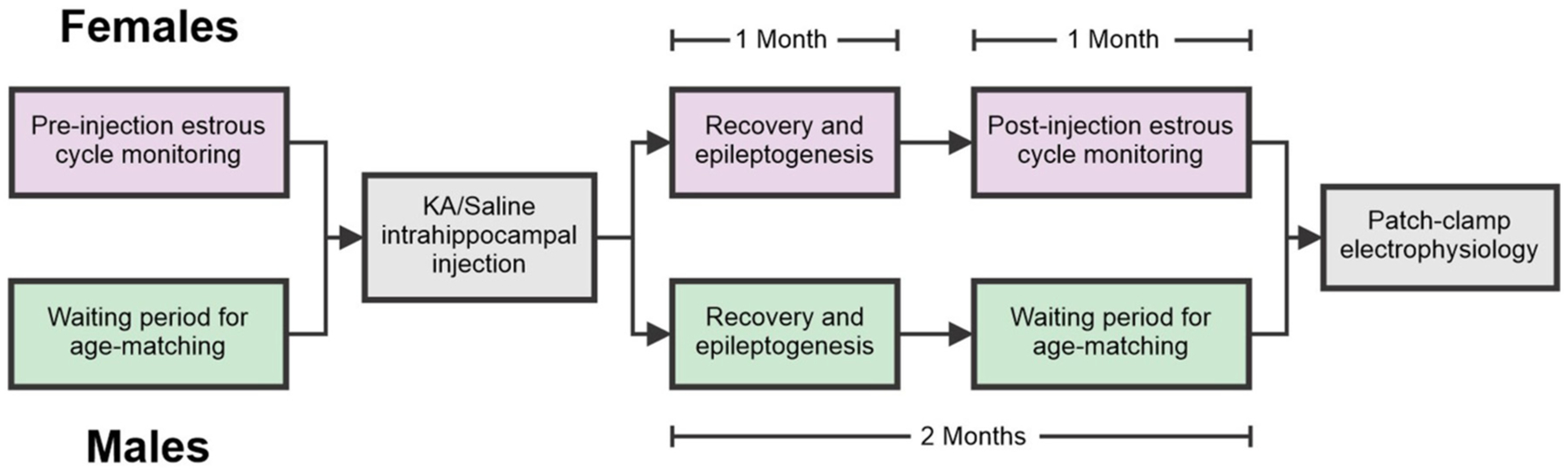
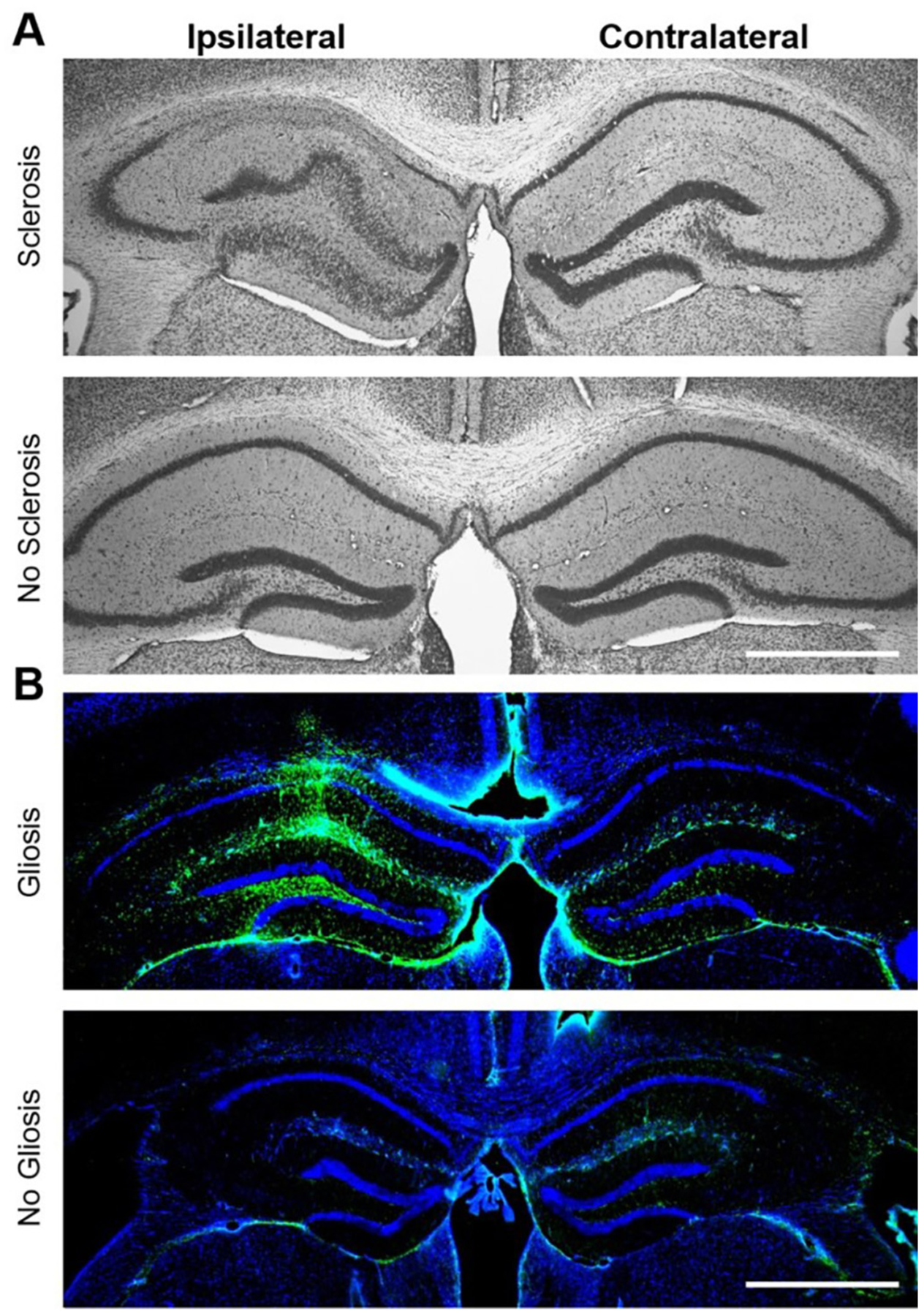
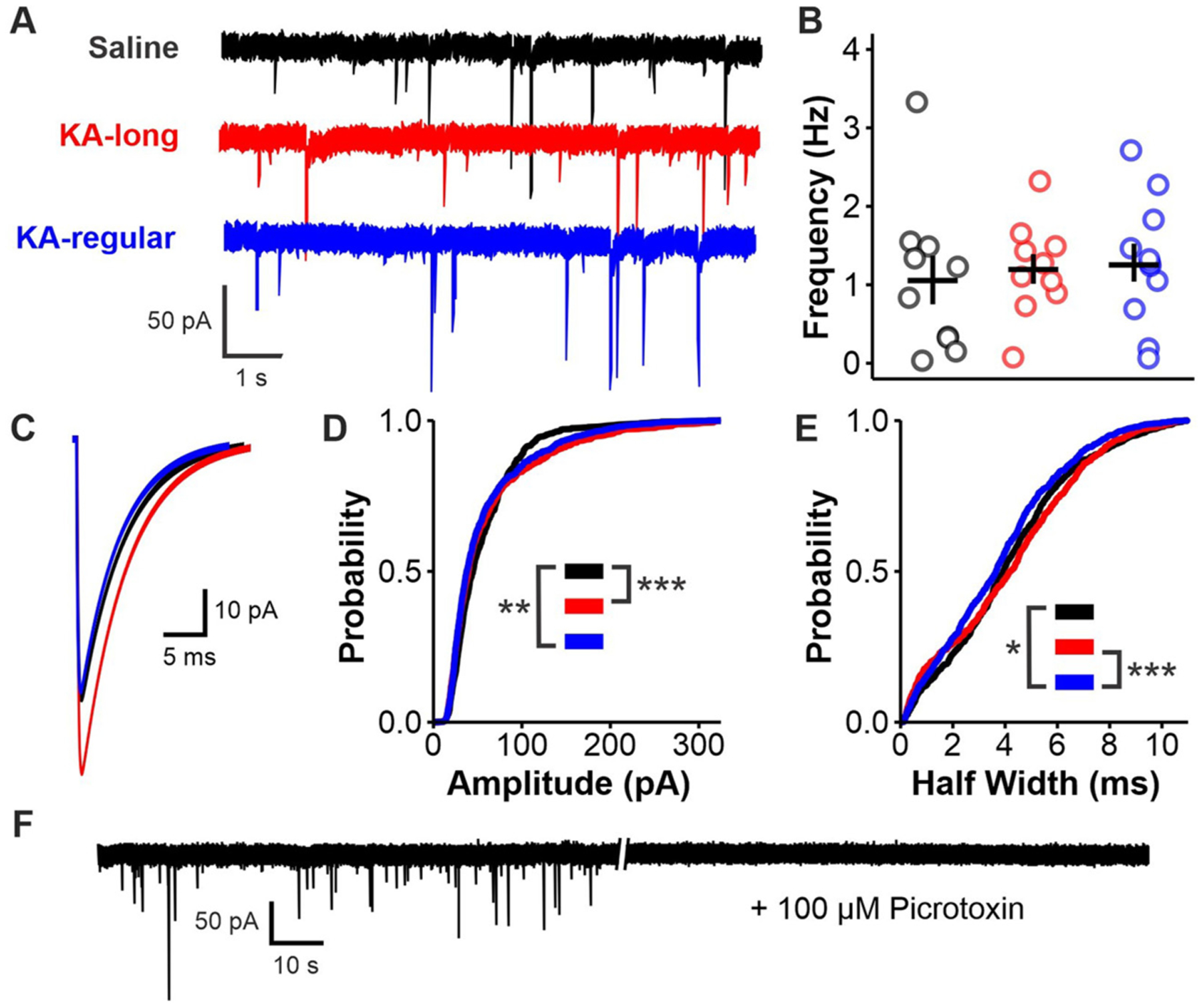
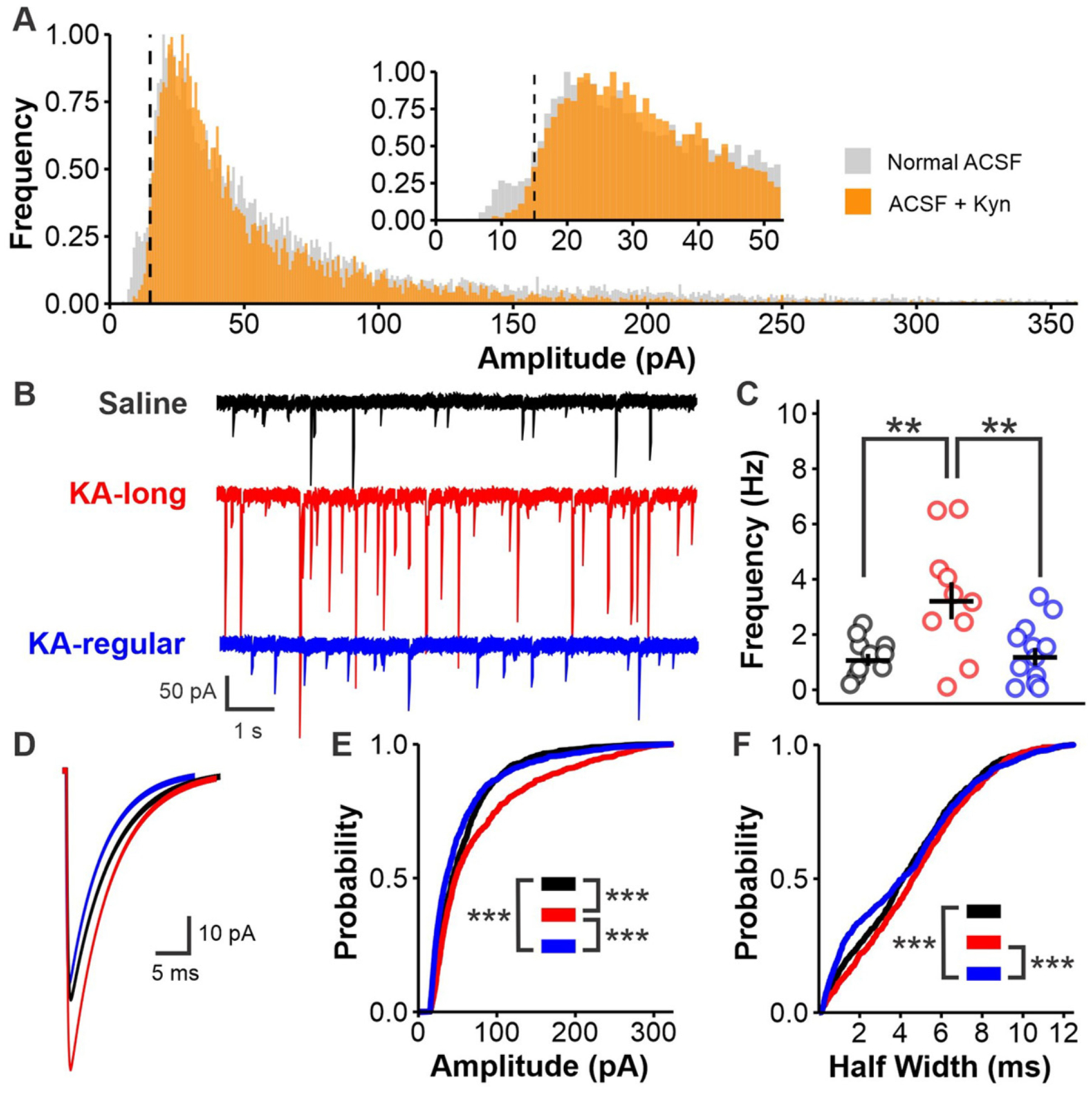
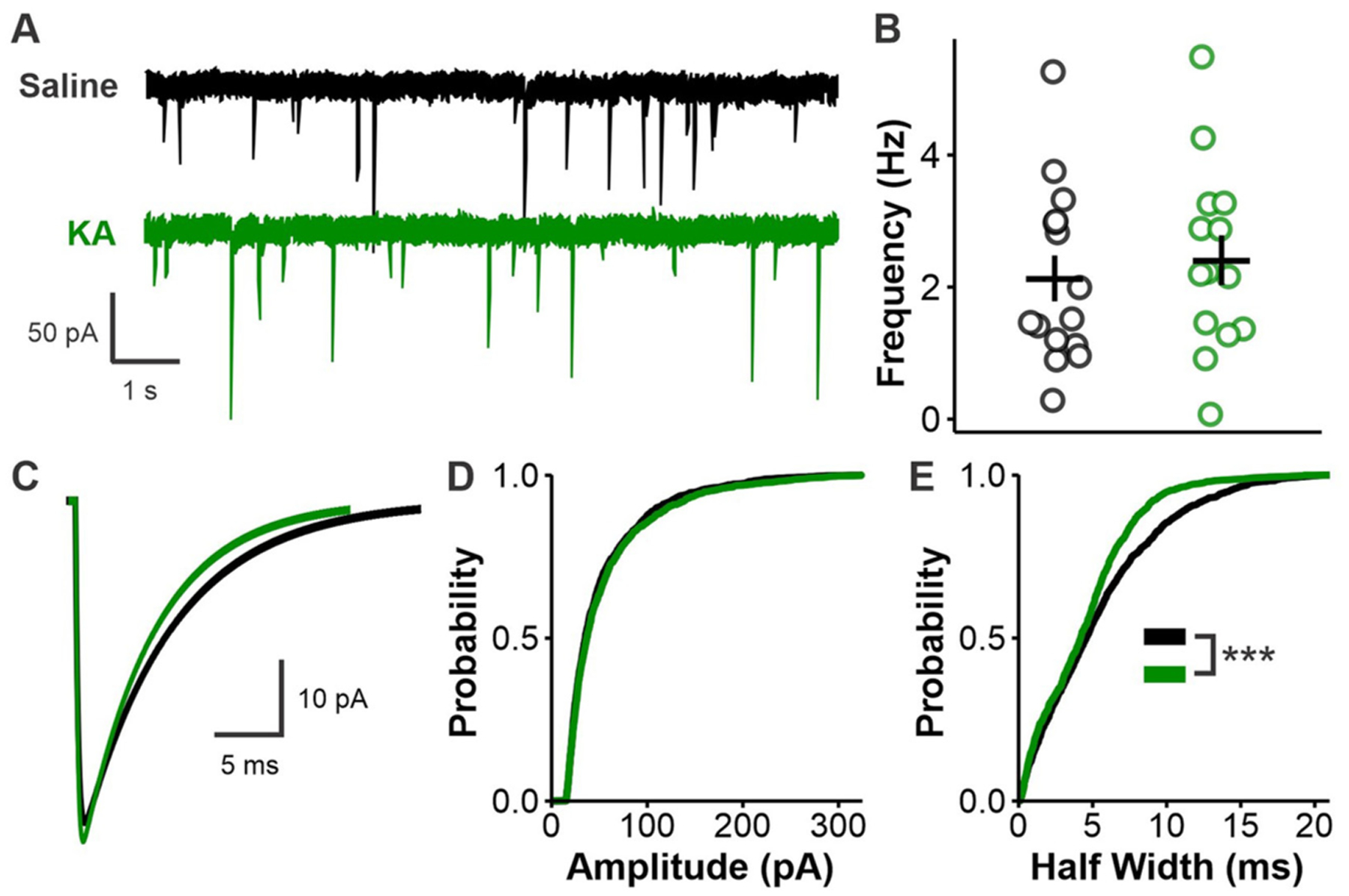
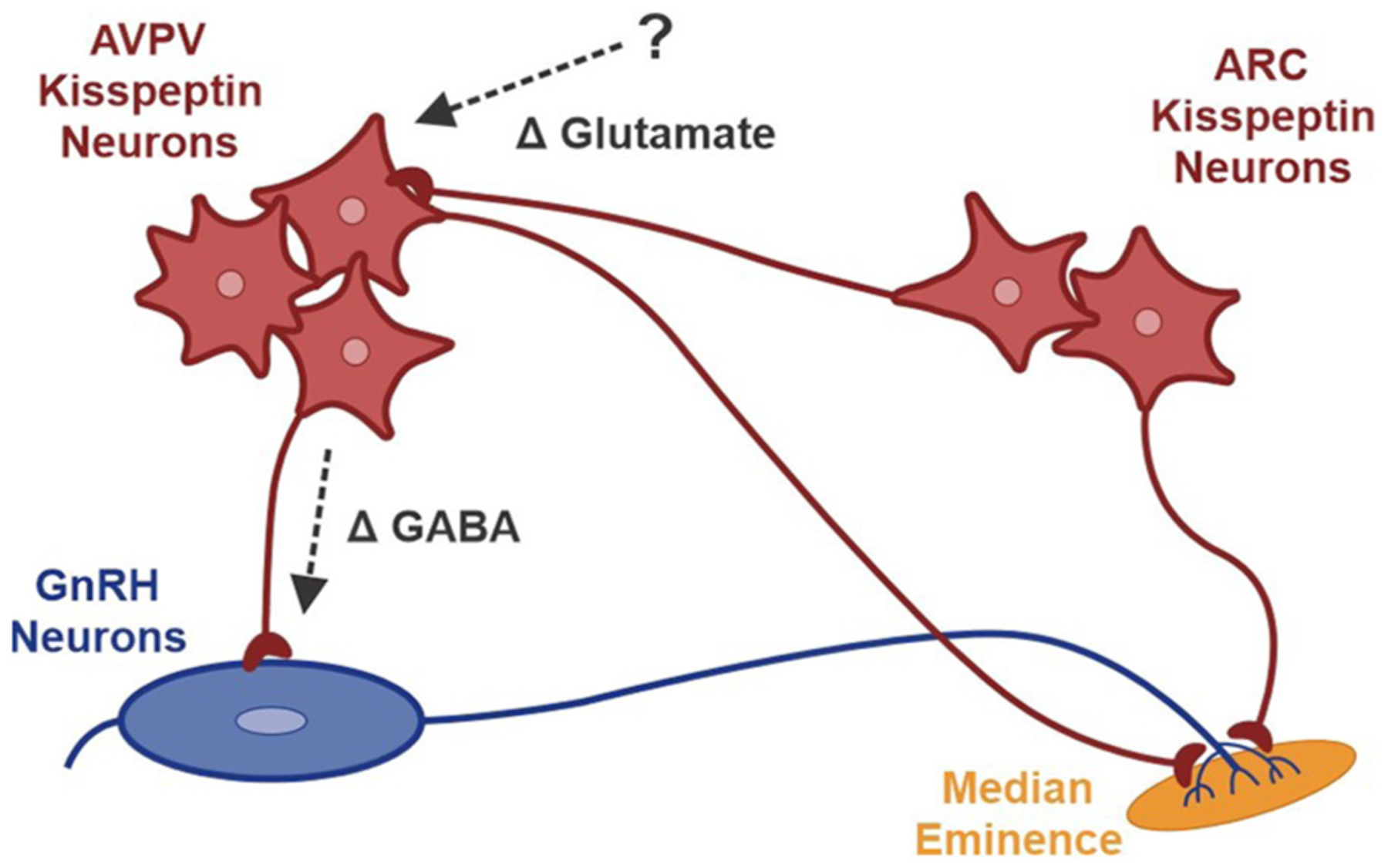
Patients with epilepsy develop reproductive endocrine comorbidities at a rate higher than that of the general population. Clinical studies have identified disrupted luteinizing hormone (LH) release patterns in patients of both sexes, suggesting potential epilepsy-associated changes in hypothalamic gonadotropin-releasing hormone (GnRH) neuron function. In previous work, we found that GnRH neuron firing is increased in diestrous females and males in the intrahippocampal kainic acid (IHKA) mouse model of temporal lobe epilepsy. Notably, GABAA receptor activation is depolarizing in adult GnRH neurons. Therefore, here we tested the hypothesis that increased GnRH neuron firing in IHKA mice is associated with increased GABAergic drive to GnRH neurons. When ionotropic glutamate receptors (iGluRs) were blocked to isolate GABAergic postsynaptic currents (PSCs), no differences in PSC frequency were seen between GnRH neurons from control and IHKA diestrous females. In the absence of iGluR blockade, however, GABA PSC frequency was increased in GnRH neurons from IHKA females with disrupted estrous cycles, but not saline-injected controls nor IHKA females without estrous cycle disruption. GABA PSC amplitude was also increased in IHKA females with disrupted estrous cycles. These findings suggest the presence of an iGluR-dependent increase in feed-forward GABAergic transmission to GnRH neurons specific to IHKA females with comorbid cycle disruption. In males, GABA PSC frequency and amplitude were unchanged but PSC duration was reduced. Together, these findings suggest that increased GABA transmission helps drive elevated firing in IHKA females on diestrus and indicate the presence of a sex-specific hypothalamic mechanism underlying reproductive endocrine dysfunction in IHKA mice.
Keywords: Electrophysiology; Epilepsy; GABA; Gonadotropin-releasing hormone; Hypothalamus; Neuroendocrinology.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest
The authors declare no competing financial interests.
Figures
Comment in
-
Sex on the Brain: Reproductive Comorbidities in Temporal Lobe Epilepsy.Epilepsy Curr. 2022 Nov 29;23(1):58-60. doi: 10.1177/15357597221135717. eCollection 2023 Jan-Feb. Epilepsy Curr. 2022. PMID: 36923330 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
