

A phylogenetically-restricted essential cell cycle progression factor in the human pathogen Candida albicans
- PMID: 35869076
- PMCID: PMC9307598
- DOI: 10.1038/s41467-022-31980-3
A phylogenetically-restricted essential cell cycle progression factor in the human pathogen Candida albicans
Abstract
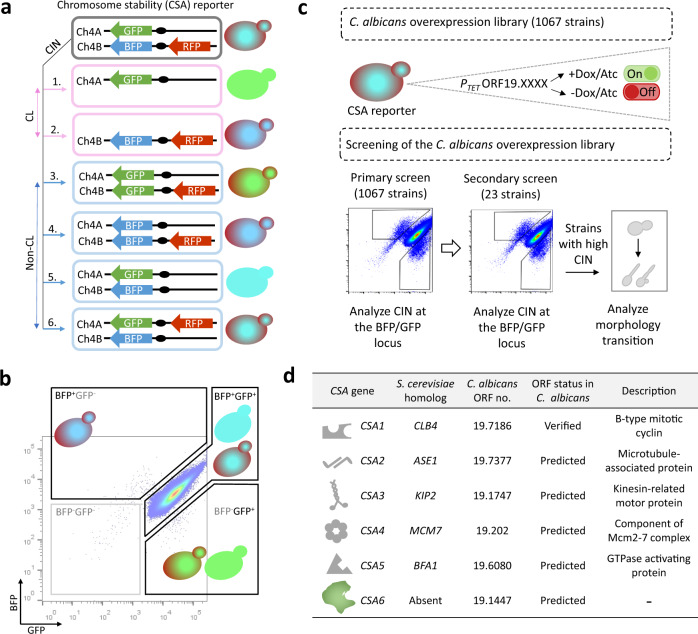
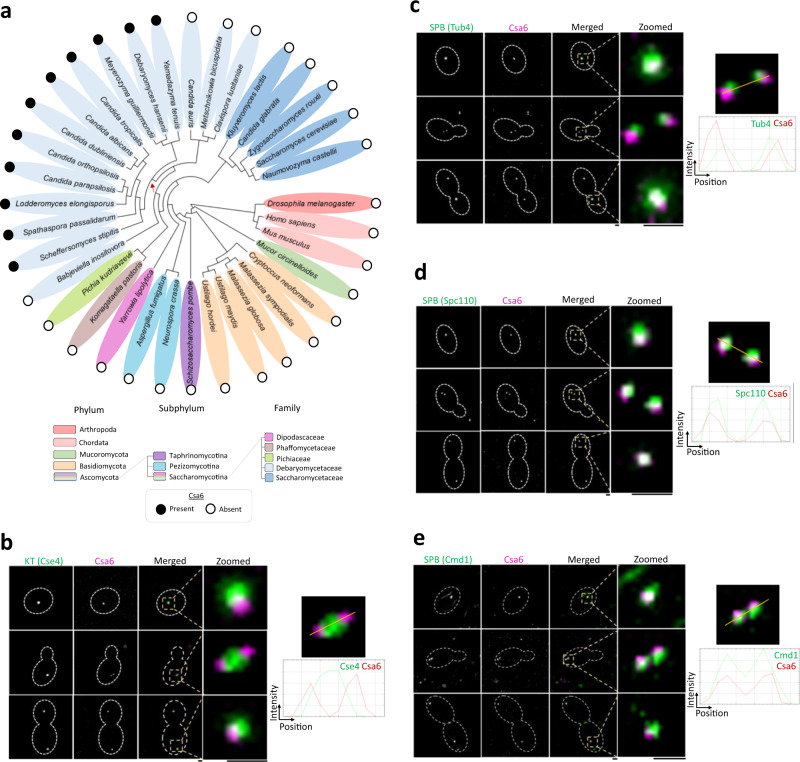
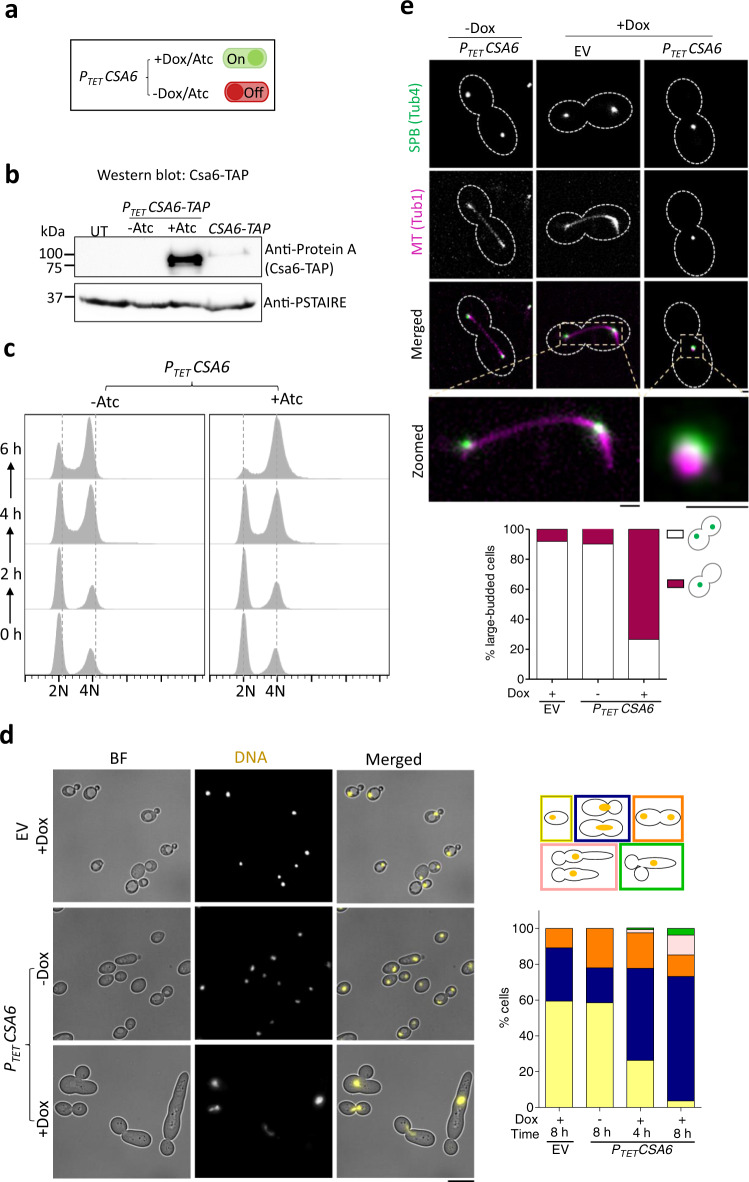
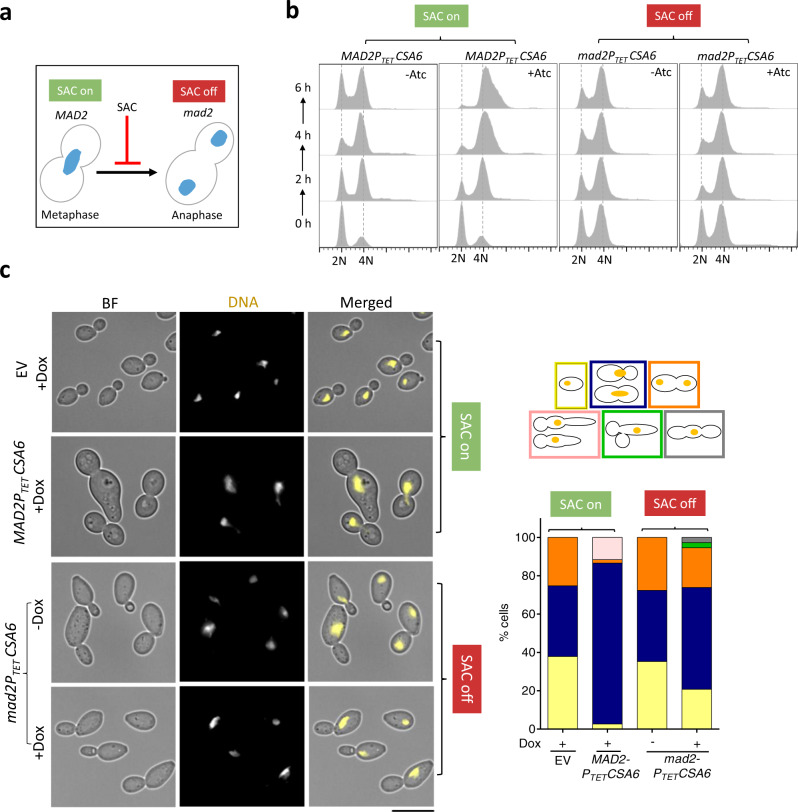
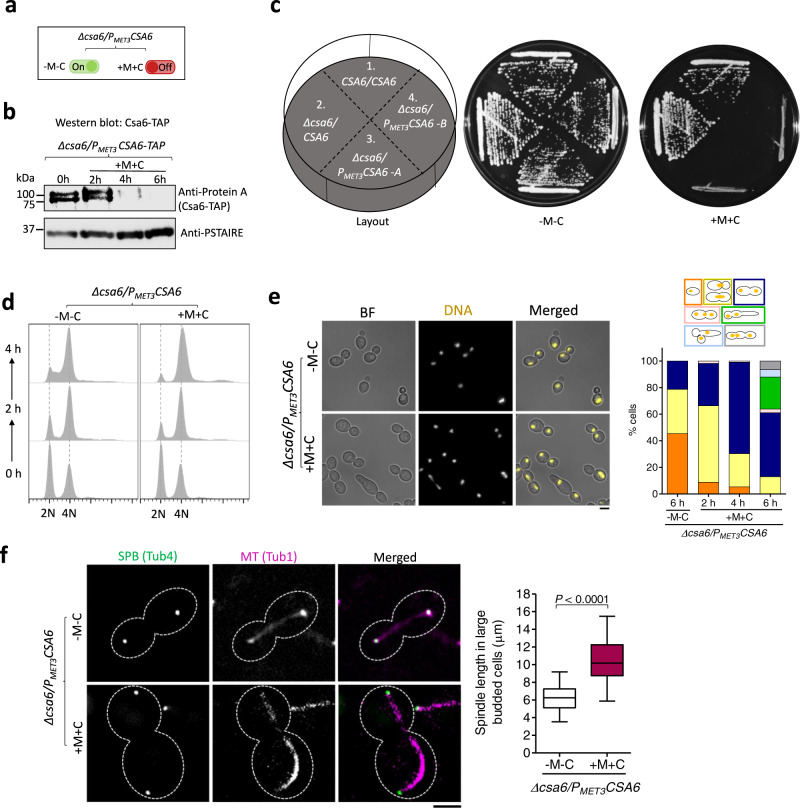
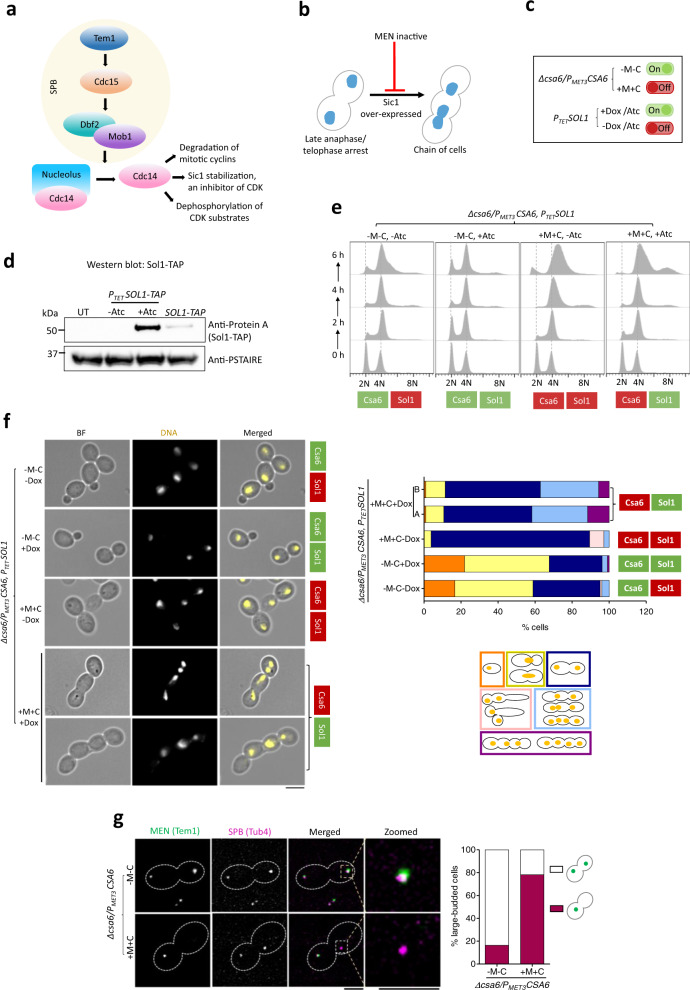
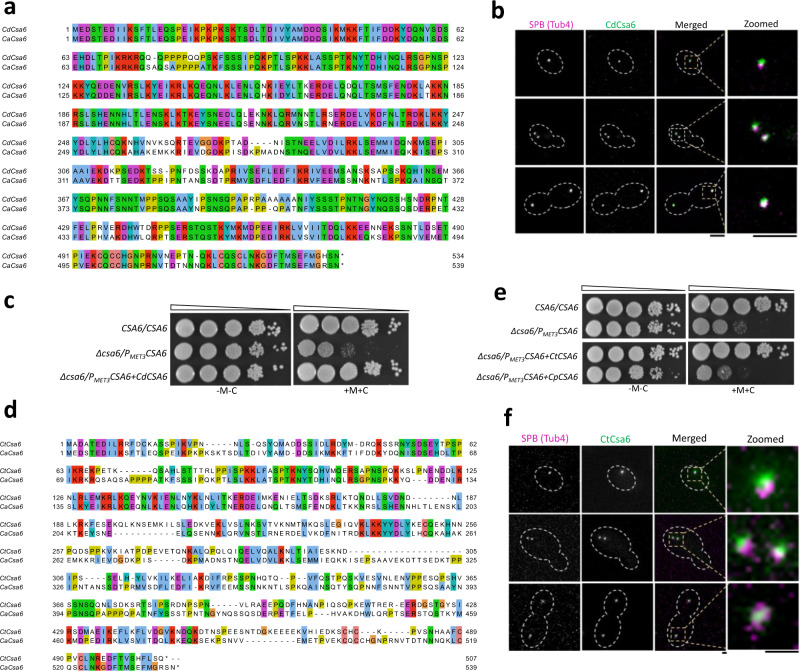
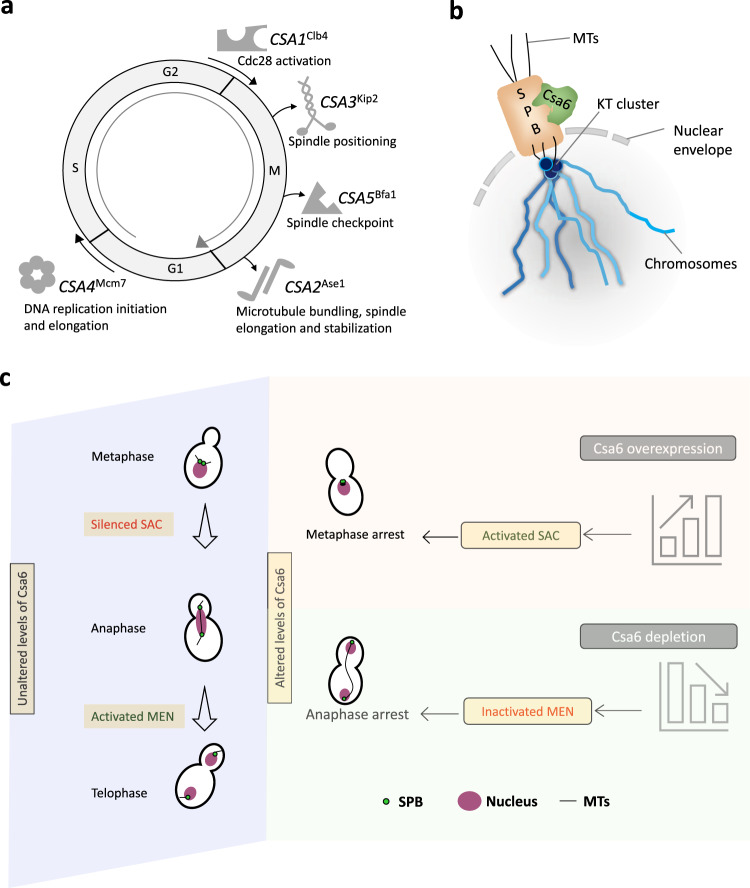
Chromosomal instability caused by cell division errors is associated with antifungal drug resistance in fungal pathogens. Here, we identify potential mechanisms underlying such instability by conducting an overexpression screen monitoring chromosomal stability in the human fungal pathogen Candida albicans. Analysis of ~1000 genes uncovers six chromosomal stability (CSA) genes, five of which are related to cell division genes of other organisms. The sixth gene, CSA6, appears to be present only in species belonging to the CUG-Ser clade, which includes C. albicans and other human fungal pathogens. The protein encoded by CSA6 localizes to the spindle pole bodies, is required for exit from mitosis, and induces a checkpoint-dependent metaphase arrest upon overexpression. Thus, Csa6 is an essential cell cycle progression factor that is restricted to the CUG-Ser fungal clade, and could therefore be explored as a potential antifungal target.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
