

A pore-forming protein drives macropinocytosis to facilitate toad water maintaining
- PMID: 35869260
- PMCID: PMC9307623
- DOI: 10.1038/s42003-022-03686-1
A pore-forming protein drives macropinocytosis to facilitate toad water maintaining
Abstract
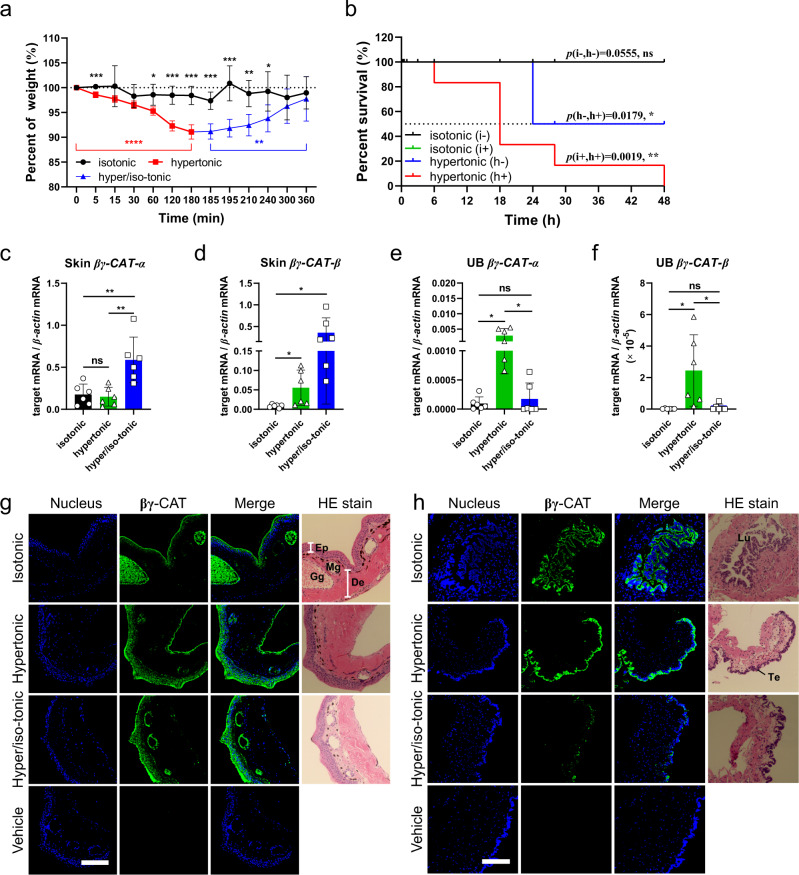
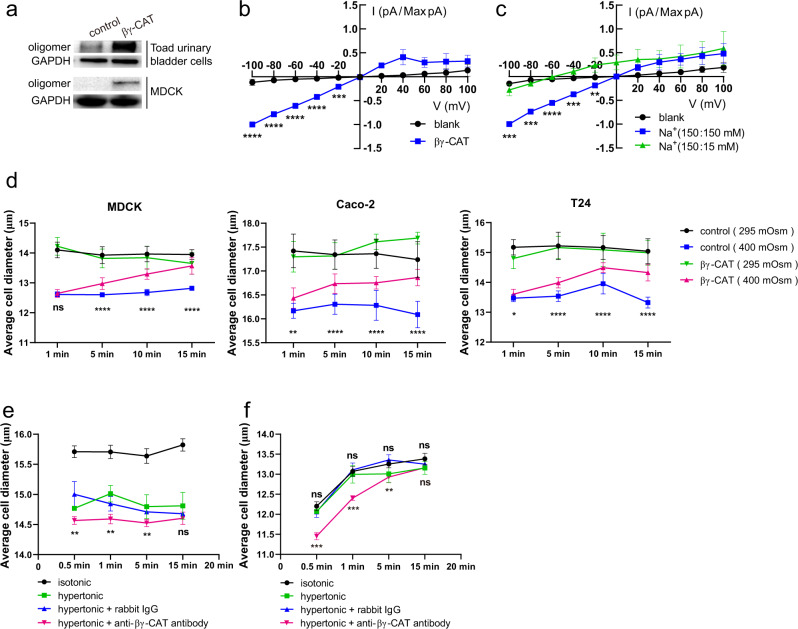
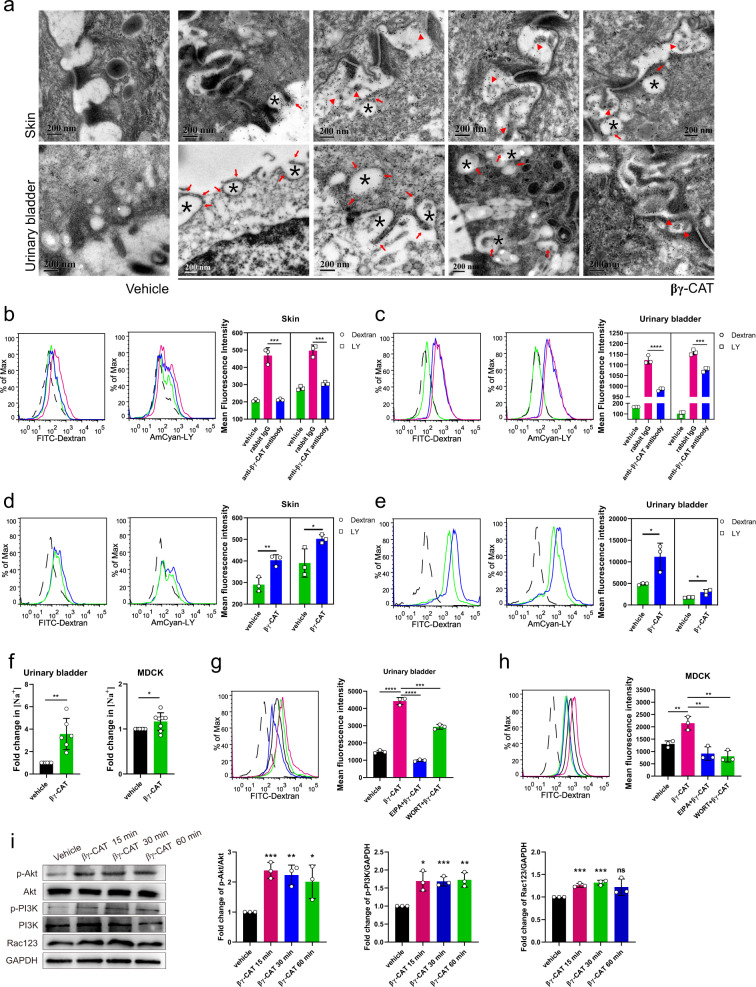
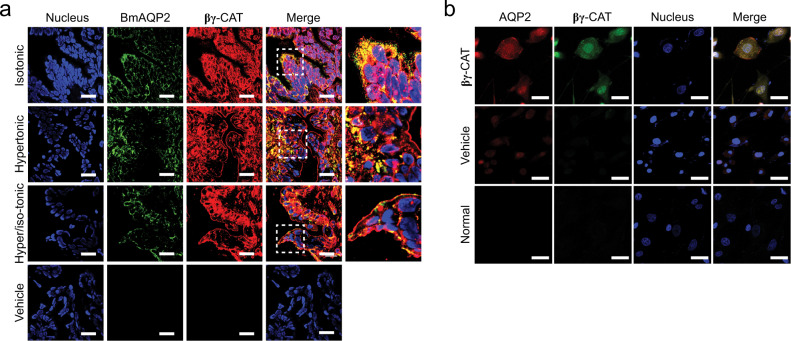
Maintaining water balance is a real challenge for amphibians in terrestrial environments. Our previous studies with toad Bombina maxima discovered a pore-forming protein and trefoil factor complex βγ-CAT, which is assembled under tight regulation depending on environmental cues. Here we report an unexpected role for βγ-CAT in toad water maintaining. Deletion of toad skin secretions, in which βγ-CAT is a major component, increased animal mortality under hypertonic stress. βγ-CAT was constitutively expressed in toad osmoregulatory organs, which was inducible under the variation of osmotic conditions. The protein induced and participated in macropinocytosis in vivo and in vitro. During extracellular hyperosmosis, βγ-CAT stimulated macropinocytosis to facilitate water import and enhanced exosomes release, which simultaneously regulated aquaporins distribution. Collectively, these findings uncovered that besides membrane integrated aquaporin, a secretory pore-forming protein can facilitate toad water maintaining via macropinocytosis induction and exocytosis modulation, especially in responses to osmotic stress.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Pore-forming protein βγ-CAT promptly responses to fasting with capacity to deliver macromolecular nutrients.FASEB J. 2022 Oct;36(10):e22533. doi: 10.1096/fj.202200528R. FASEB J. 2022. PMID: 36065711
-
IgG Fc-binding protein positively regulates the assembly of pore-forming protein complex βγ-CAT evolved to drive cell vesicular delivery and transport.J Biol Chem. 2023 Jun;299(6):104717. doi: 10.1016/j.jbc.2023.104717. Epub 2023 Apr 15. J Biol Chem. 2023. PMID: 37068610 Free PMC article.
-
A Pore Forming Toxin-like Protein Derived from Chinese Red Belly Toad Bombina maxima Triggers the Pyroptosis of Hippomal Neural Cells and Impairs the Cognitive Ability of Mice.Toxins (Basel). 2023 Mar 3;15(3):191. doi: 10.3390/toxins15030191. Toxins (Basel). 2023. PMID: 36977082 Free PMC article.
-
Animal secretory endolysosome channel discovery.Zool Res. 2021 Mar 18;42(2):141-152. doi: 10.24272/j.issn.2095-8137.2020.358. Zool Res. 2021. PMID: 33527802 Free PMC article. Review.
-
Molecular diversity of vasotocin-dependent aquaporins closely associated with water adaptation strategy in anuran amphibians.J Neuroendocrinol. 2010 May;22(5):407-12. doi: 10.1111/j.1365-2826.2010.01981.x. Epub 2010 Feb 12. J Neuroendocrinol. 2010. PMID: 20163517 Review.
Cited by
-
The Bi-Functional Paxilline Enriched in Skin Secretion of Tree Frogs (Hyla japonica) Targets the KCNK18 and BKCa Channels.Toxins (Basel). 2023 Jan 12;15(1):70. doi: 10.3390/toxins15010070. Toxins (Basel). 2023. PMID: 36668889 Free PMC article.
-
Amphibian pore-forming protein βγ-CAT drives extracellular nutrient scavenging under cell nutrient deficiency.iScience. 2023 Apr 7;26(5):106598. doi: 10.1016/j.isci.2023.106598. eCollection 2023 May 19. iScience. 2023. PMID: 37128610 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
