

Sex-biased and parental allele-specific gene regulation by KDM6A
- PMID: 35871105
- PMCID: PMC9308343
- DOI: 10.1186/s13293-022-00452-0
Sex-biased and parental allele-specific gene regulation by KDM6A
Abstract
Background: KDM6A is a demethylase encoded by a gene with female-biased expression due to escape from X inactivation. Its main role is to facilitate gene expression through removal of the repressive H3K27me3 mark, with evidence of some additional histone demethylase-independent functions. KDM6A mutations have been implicated in congenital disorders such as Kabuki Syndrome, as well as in sex differences in cancer.
Methods: Kdm6a was knocked out using CRISPR/Cas9 gene editing in F1 male and female mouse embryonic stem cells (ES) derived from reciprocal crosses between C57BL6 x Mus castaneus. Diploid and allelic RNA-seq analyses were done to compare gene expression between wild-type and Kdm6a knockout (KO) clones. The effects of Kdm6a KO on sex-biased gene expression were investigated by comparing gene expression between male and female ES cells. Changes in H3K27me3 enrichment and chromatin accessibility at promoter regions of genes with expression changes were characterized by ChIP-seq and ATAC-seq followed by diploid and allelic analyses.
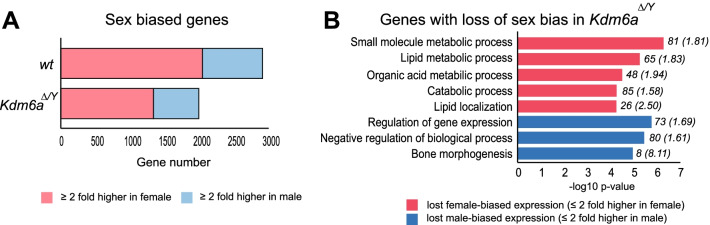
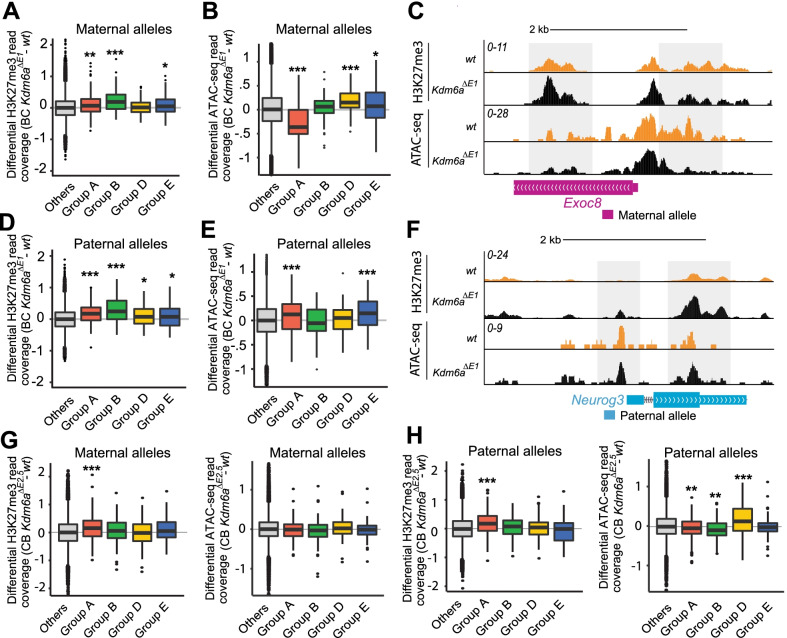
Results: We report that Kdm6a KO in male and female embryonic stem (ES) cells derived from F1 hybrid mice cause extensive gene dysregulation, disruption of sex biases, and specific parental allele effects. Among the dysregulated genes are candidate genes that may explain abnormal developmental features of Kabuki syndrome caused by KDM6A mutations in human. Strikingly, Kdm6a knockouts result in a decrease in sex-biased expression and in preferential downregulation of the maternal alleles of a number of genes. Most promoters of dysregulated genes show concordant epigenetic changes including gain of H3K27me3 and loss of chromatin accessibility, but there was less concordance when considering allelic changes.
Conclusions: Our study reveals new sex-related roles of KDM6A in the regulation of developmental genes, the maintenance of sex-biased gene expression, and the differential expression of parental alleles.
Keywords: Allele-specific; Chromatin; Development; Epigenetics; Gene regulation; Histone methylation; Imprinting; Parent-of-origin; Sex biases.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
