

Transcription factor CmbHLH16 regulates petal anthocyanin homeostasis under different lights in Chrysanthemum
- PMID: 35876821
- PMCID: PMC9516746
- DOI: 10.1093/plphys/kiac342
Transcription factor CmbHLH16 regulates petal anthocyanin homeostasis under different lights in Chrysanthemum
Abstract
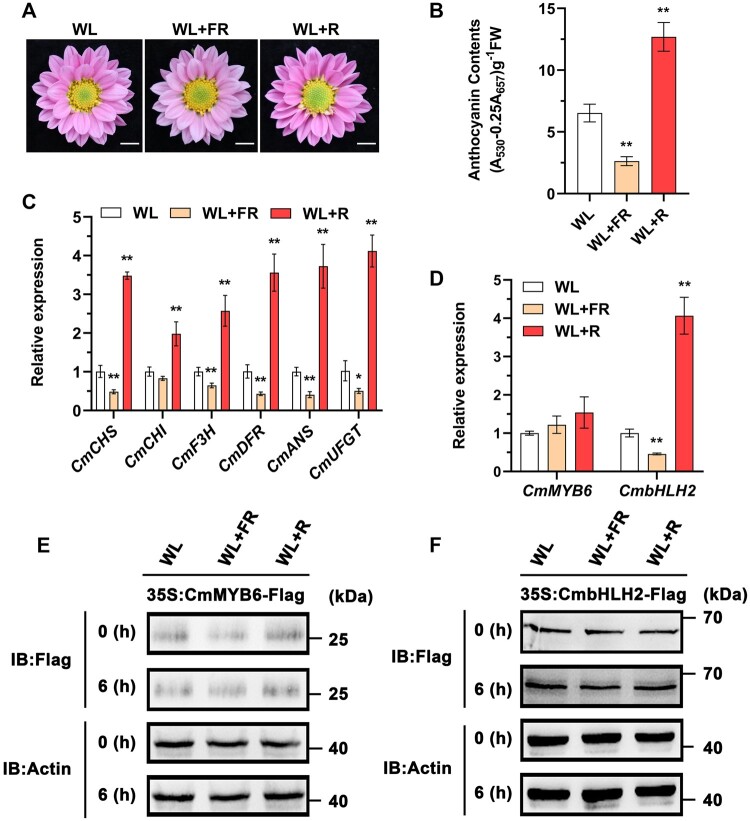
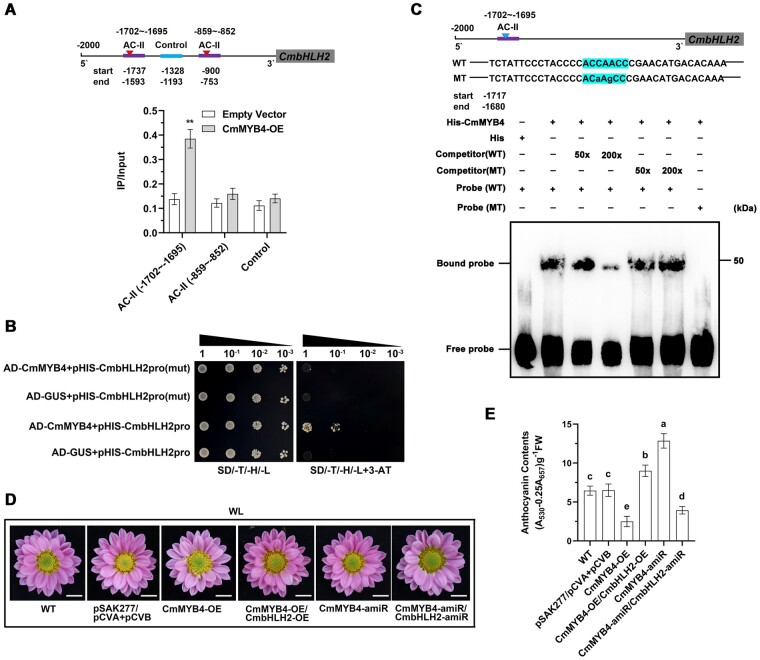
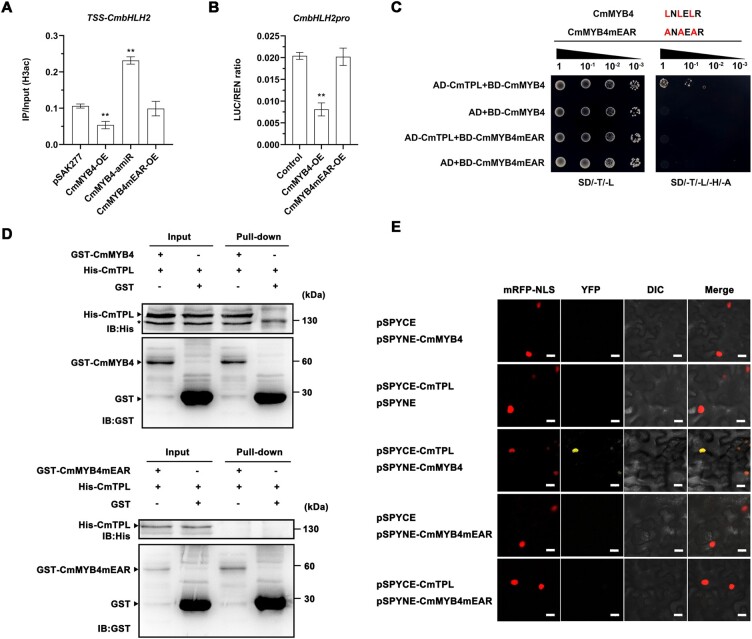
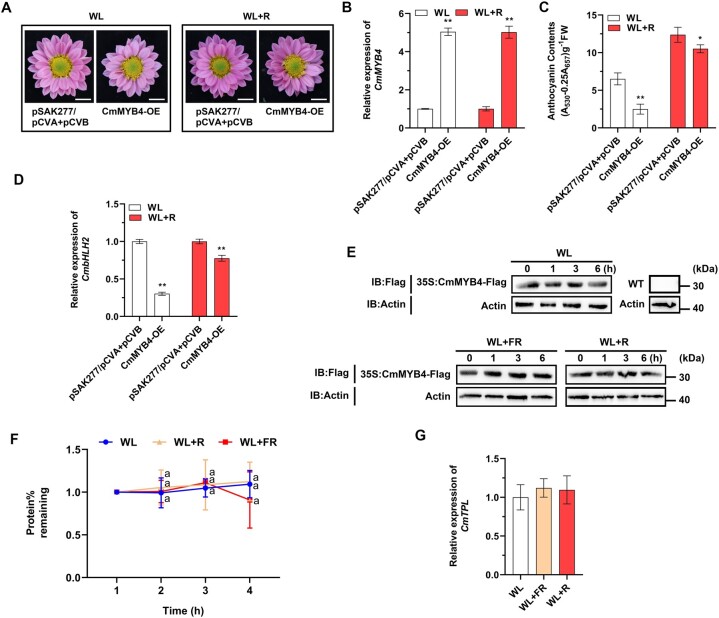
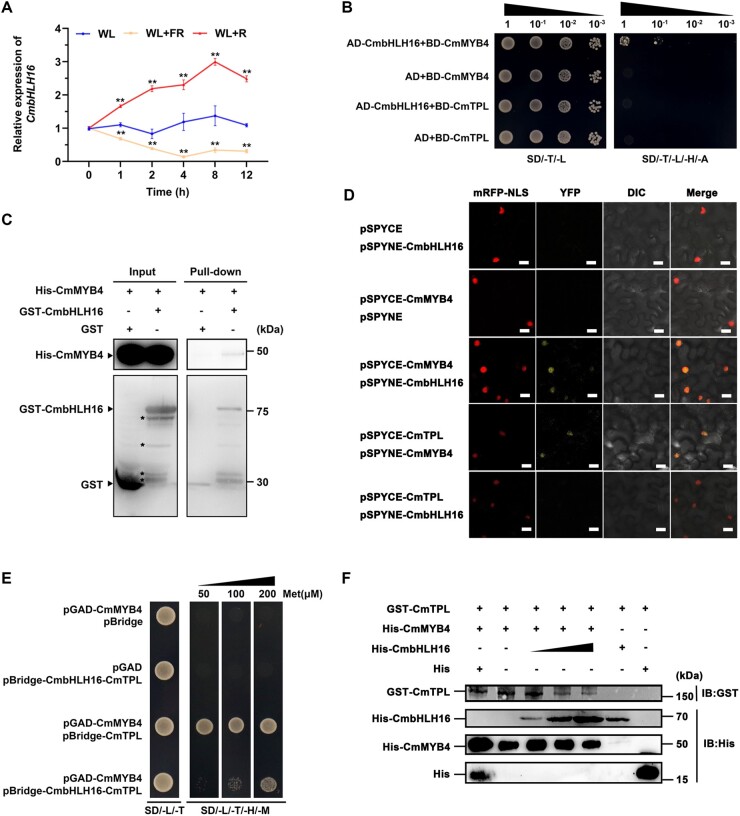
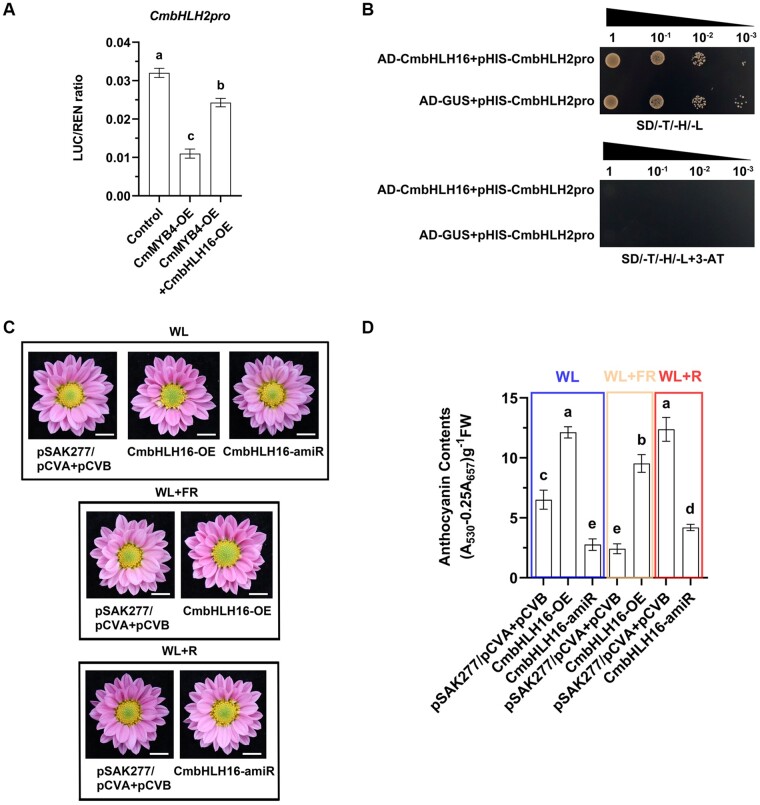
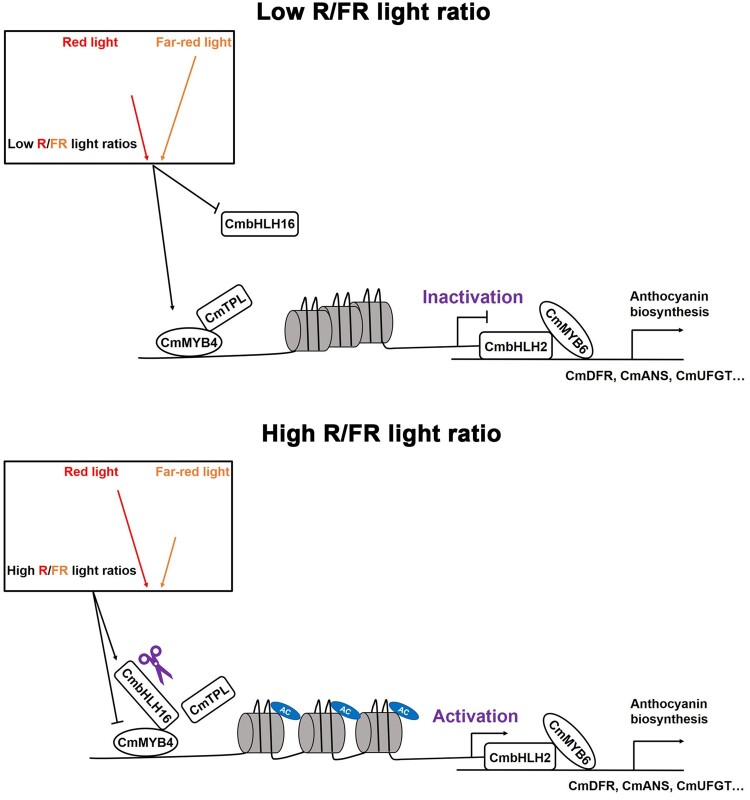
Light is essential to plant survival and elicits a wide range of plant developmental and physiological responses under different light conditions. A low red-to-far red (R/FR) light ratio induces shade-avoidance responses, including decreased anthocyanin accumulation, whereas a high R/FR light ratio promotes anthocyanin biosynthesis. However, the detailed molecular mechanism underpinning how different R/FR light ratios regulate anthocyanin homeostasis remains elusive, especially in non-model species. Here, we demonstrate that a low R/FR light ratio induced the expression of CmMYB4, which suppressed the anthocyanin activator complex CmMYB6-CmbHLH2, leading to the reduction of anthocyanin accumulation in Chrysanthemum (Chrysanthemum morifolium) petals. Specifically, CmMYB4 recruited the corepressor CmTPL (TOPLESS) to directly bind the CmbHLH2 promoter and suppressed its transcription by impairing histone H3 acetylation. Moreover, the low R/FR light ratio inhibited the PHYTOCHROME INTERACTING FACTOR family transcription factor CmbHLH16, which can competitively bind to CmMYB4 and destabilize the CmMYB4-CmTPL protein complex. Under the high R/FR light ratio, CmbHLH16 was upregulated, which impeded the formation of the CmMYB4-CmTPL complex and released the suppression of CmbHLH2, thus promoting anthocyanin accumulation in Chrysanthemum petals. Our findings reveal a mechanism by which different R/FR light ratios fine-tune anthocyanin homeostasis in flower petals.
© American Society of Plant Biologists 2022. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures
Similar articles
-
CmNAC25 targets CmMYB6 to positively regulate anthocyanin biosynthesis during the post-flowering stage in chrysanthemum.BMC Biol. 2023 Oct 9;21(1):211. doi: 10.1186/s12915-023-01719-7. BMC Biol. 2023. PMID: 37807042 Free PMC article.
-
Ectopic Expression of Multiple Chrysanthemum (Chrysanthemum × morifolium) R2R3-MYB Transcription Factor Genes Regulates Anthocyanin Accumulation in Tobacco.Genes (Basel). 2019 Oct 4;10(10):777. doi: 10.3390/genes10100777. Genes (Basel). 2019. PMID: 31590246 Free PMC article.
-
A Novel bHLH Transcription Factor Involved in Regulating Anthocyanin Biosynthesis in Chrysanthemums (Chrysanthemum morifolium Ramat.).PLoS One. 2015 Nov 30;10(11):e0143892. doi: 10.1371/journal.pone.0143892. eCollection 2015. PLoS One. 2015. PMID: 26619181 Free PMC article.
-
Anthocyanins in Floral Colors: Biosynthesis and Regulation in Chrysanthemum Flowers.Int J Mol Sci. 2020 Sep 7;21(18):6537. doi: 10.3390/ijms21186537. Int J Mol Sci. 2020. PMID: 32906764 Free PMC article. Review.
-
Achievements and perspectives in biochemistry concerning anthocyanin modification for blue flower coloration.Plant Cell Physiol. 2015 Jan;56(1):28-40. doi: 10.1093/pcp/pcu097. Epub 2014 Jul 10. Plant Cell Physiol. 2015. PMID: 25015943 Review.
Cited by
-
The Y locus encodes a REPRESSOR OF PHOTOSYNTHETIC GENES protein that represses carotenoid biosynthesis via interaction with APRR2 in carrot.Plant Cell. 2024 Jul 31;36(8):2798-2817. doi: 10.1093/plcell/koae111. Plant Cell. 2024. PMID: 38593056 Free PMC article.
-
Transcription factors LvBBX24 and LvbZIP44 coordinated anthocyanin accumulation in response to light in lily petals.Hortic Res. 2024 Jul 30;11(10):uhae211. doi: 10.1093/hr/uhae211. eCollection 2024 Oct. Hortic Res. 2024. PMID: 39372289 Free PMC article.
-
Transcriptomic and metabolomic analyses reveal CmMYB308 as a key regulator in the pink flower color variation of 'Dante Purple' chrysanthemum.Plant Cell Rep. 2024 May 31;43(6):157. doi: 10.1007/s00299-024-03244-5. Plant Cell Rep. 2024. PMID: 38819475
-
Independent flavonoid and anthocyanin biosynthesis in the flesh of a red-fleshed table grape revealed by metabolome and transcriptome co-analysis.BMC Plant Biol. 2023 Jul 15;23(1):361. doi: 10.1186/s12870-023-04368-8. BMC Plant Biol. 2023. PMID: 37454071 Free PMC article.
-
CmNAC25 targets CmMYB6 to positively regulate anthocyanin biosynthesis during the post-flowering stage in chrysanthemum.BMC Biol. 2023 Oct 9;21(1):211. doi: 10.1186/s12915-023-01719-7. BMC Biol. 2023. PMID: 37807042 Free PMC article.
References
-
- Aharoni A, De Vos CR, Wein M, Sun Z, Greco R, Kroon A, Mol JN, O’Connell AP (2001) The strawberry FaMYB1 transcription factor suppresses anthocyanin and flavonol accumulation in transgenic tobacco. Plant J 28: 319–332 - PubMed
-
- Bae G, Choi G (2008) Decoding of light signals by plant phytochromes and their interacting proteins. Annu Rev Plant Biol 59: 281–311 - PubMed
-
- Ballaré CL (2009) Illuminated behaviour: phytochrome as a key regulator of light foraging and plant anti‐herbivore defence. Plant Cell Environ 32: 713–725 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
