

Molecular framework integrating nitrate sensing in root and auxin-guided shoot adaptive responses
- PMID: 35878040
- PMCID: PMC9351359
- DOI: 10.1073/pnas.2122460119
Molecular framework integrating nitrate sensing in root and auxin-guided shoot adaptive responses
Abstract
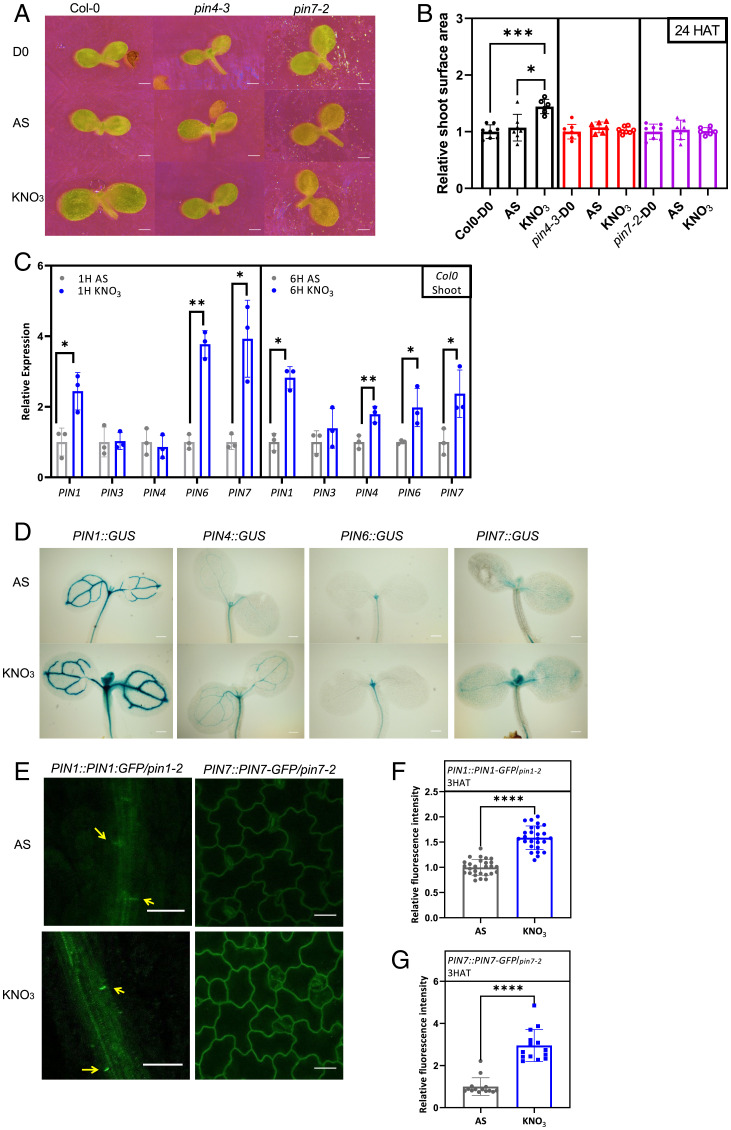
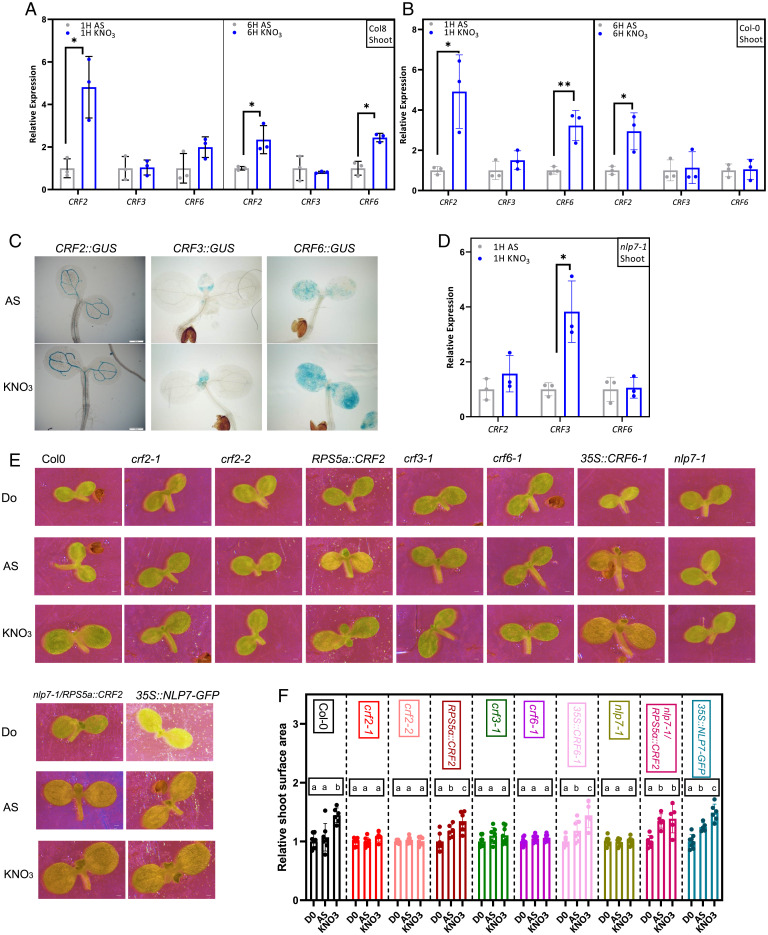
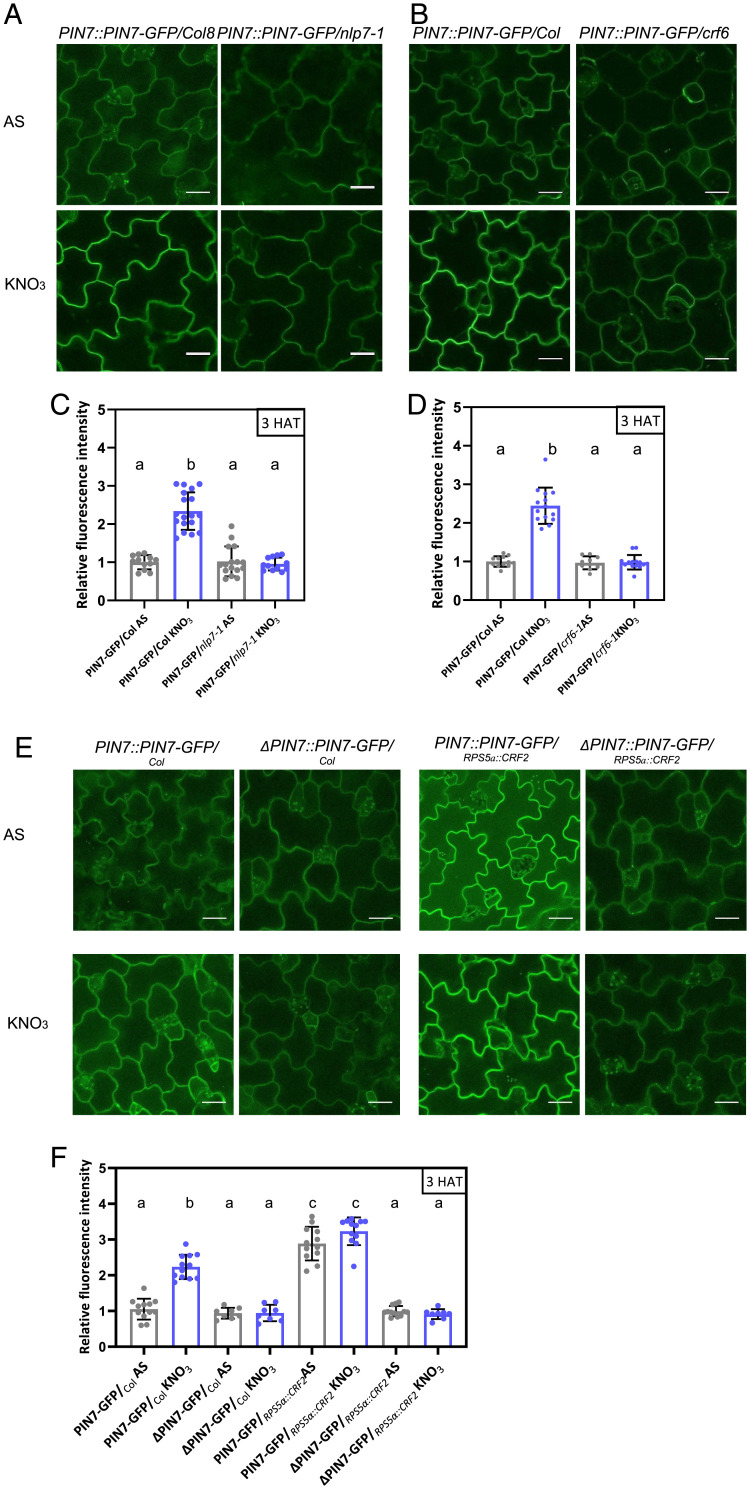
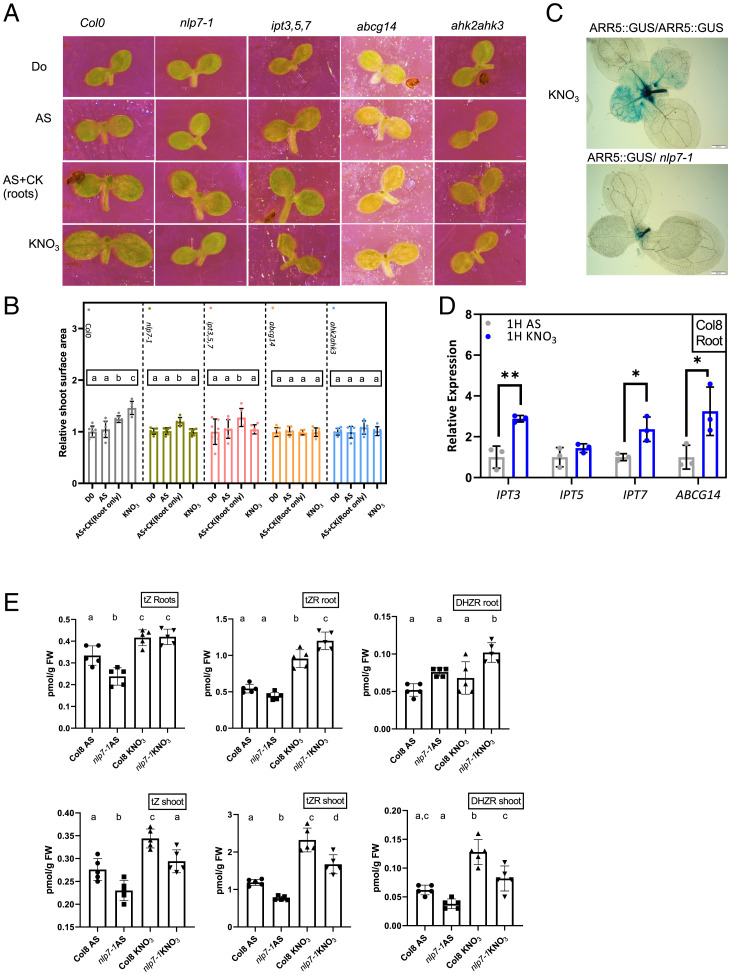
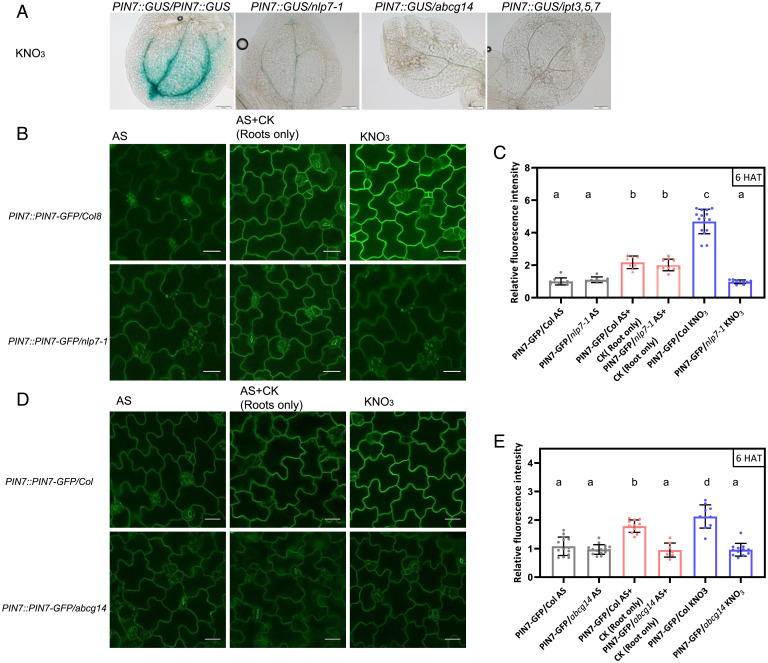
Mineral nutrition is one of the key environmental factors determining plant development and growth. Nitrate is the major form of macronutrient nitrogen that plants take up from the soil. Fluctuating availability or deficiency of this element severely limits plant growth and negatively affects crop production in the agricultural system. To cope with the heterogeneity of nitrate distribution in soil, plants evolved a complex regulatory mechanism that allows rapid adjustment of physiological and developmental processes to the status of this nutrient. The root, as a major exploitation organ that controls the uptake of nitrate to the plant body, acts as a regulatory hub that, according to nitrate availability, coordinates the growth and development of other plant organs. Here, we identified a regulatory framework, where cytokinin response factors (CRFs) play a central role as a molecular readout of the nitrate status in roots to guide shoot adaptive developmental response. We show that nitrate-driven activation of NLP7, a master regulator of nitrate response in plants, fine tunes biosynthesis of cytokinin in roots and its translocation to shoots where it enhances expression of CRFs. CRFs, through direct transcriptional regulation of PIN auxin transporters, promote the flow of auxin and thereby stimulate the development of shoot organs.
Keywords: macronutrient; nitrate; plant development.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
