

Dysfunctional ERG signaling drives pulmonary vascular aging and persistent fibrosis
- PMID: 35879310
- PMCID: PMC9314350
- DOI: 10.1038/s41467-022-31890-4
Dysfunctional ERG signaling drives pulmonary vascular aging and persistent fibrosis
Erratum in
-
Author Correction: Dysfunctional ERG signaling drives pulmonary vascular aging and persistent fibrosis.Nat Commun. 2022 Sep 28;13(1):5687. doi: 10.1038/s41467-022-33197-w. Nat Commun. 2022. PMID: 36171208 Free PMC article. No abstract available.
Abstract
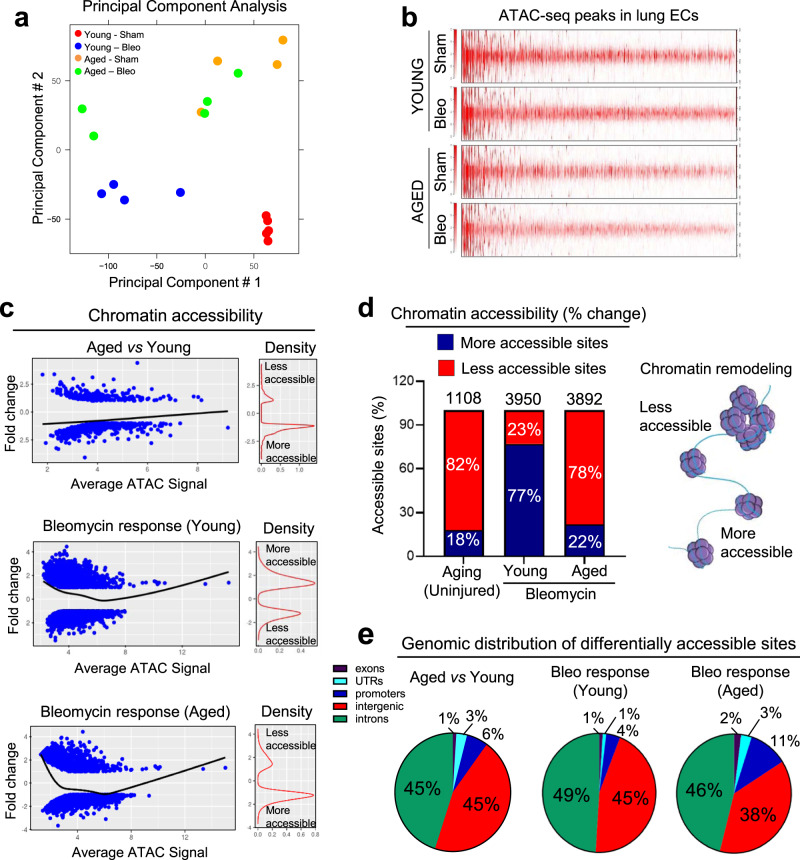
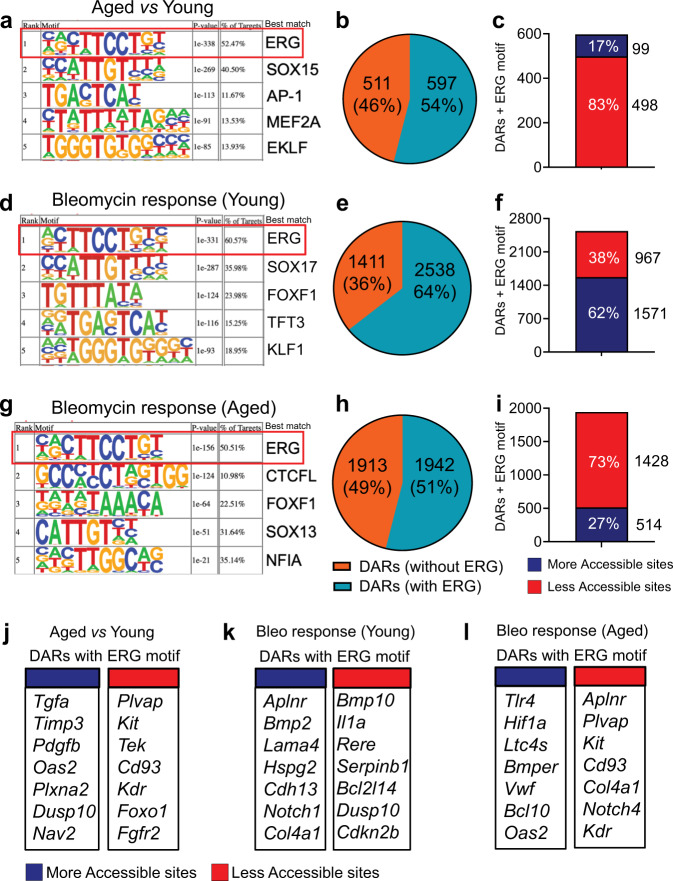
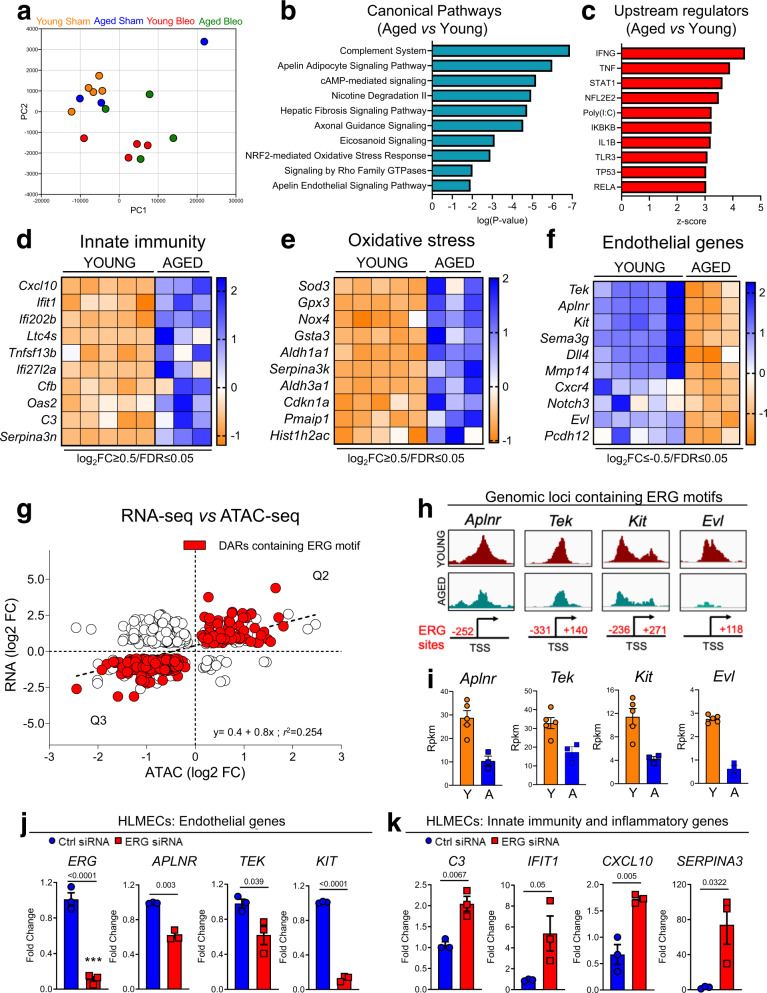
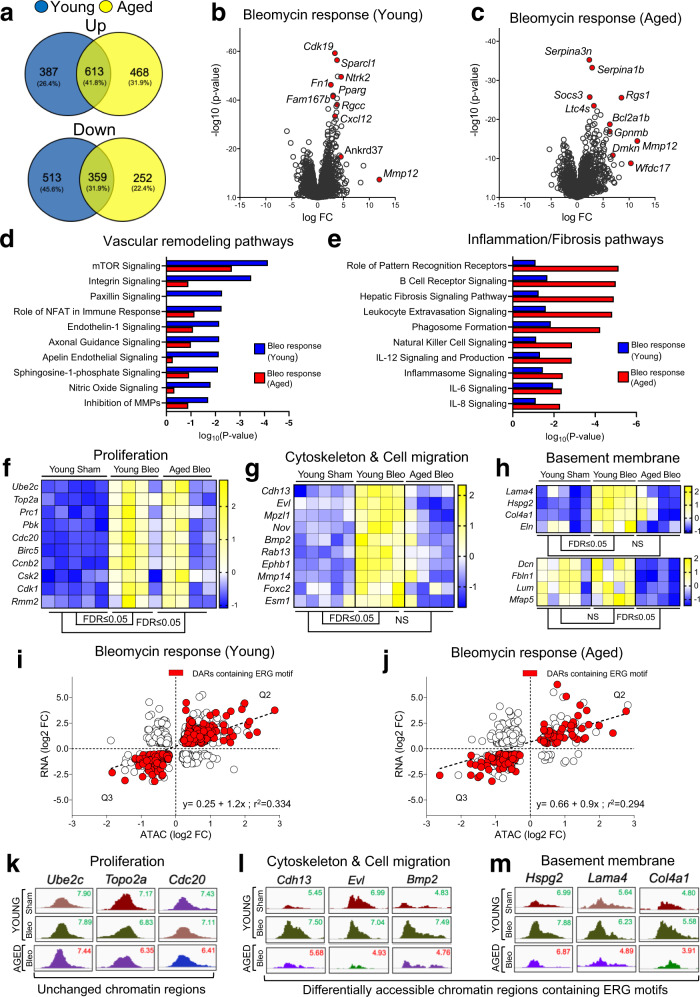
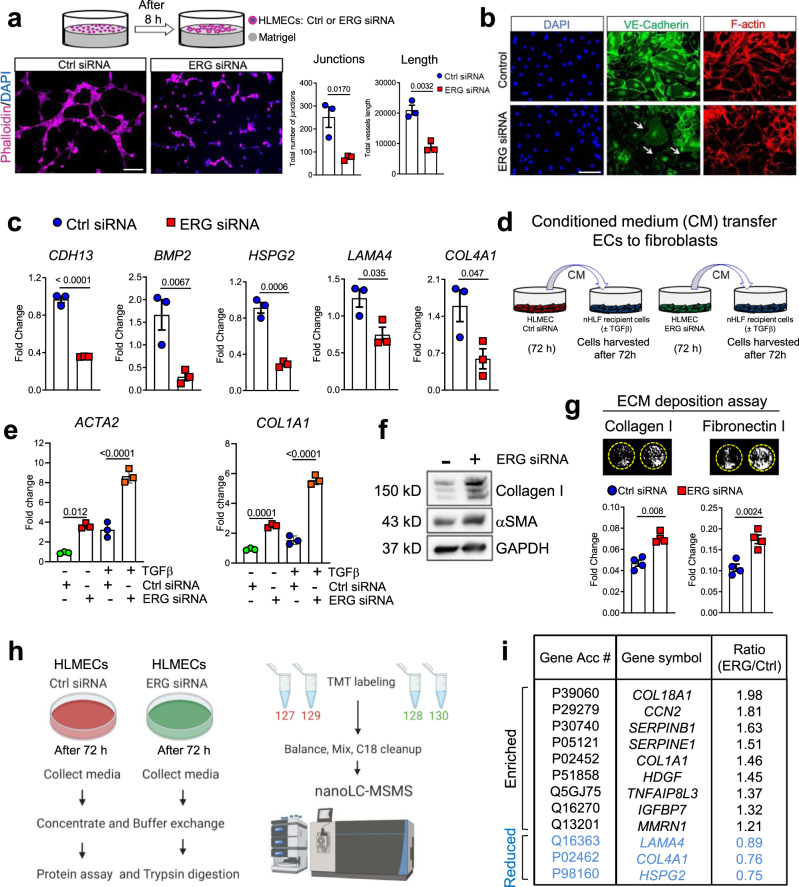
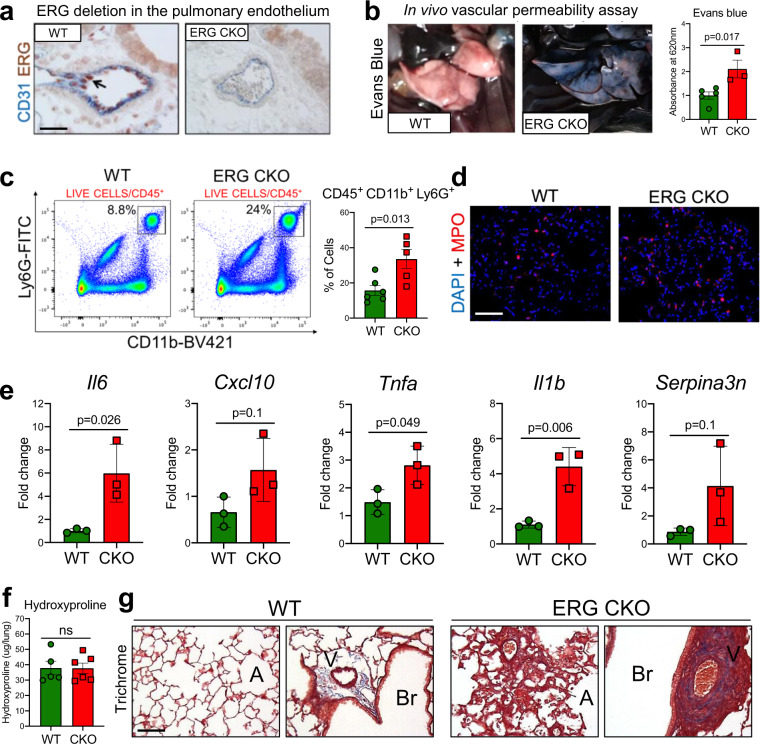
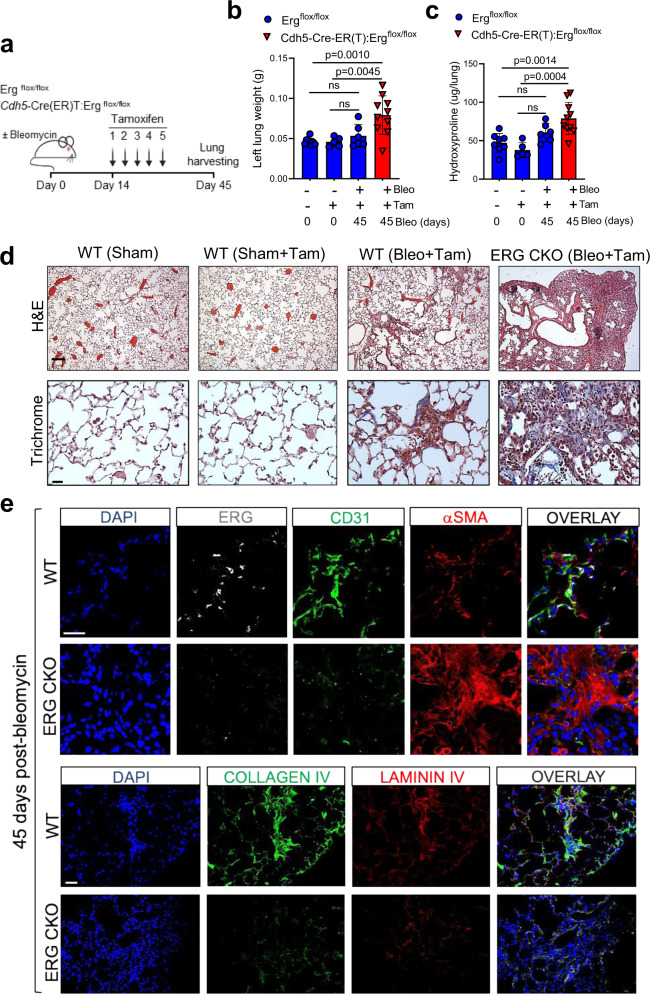
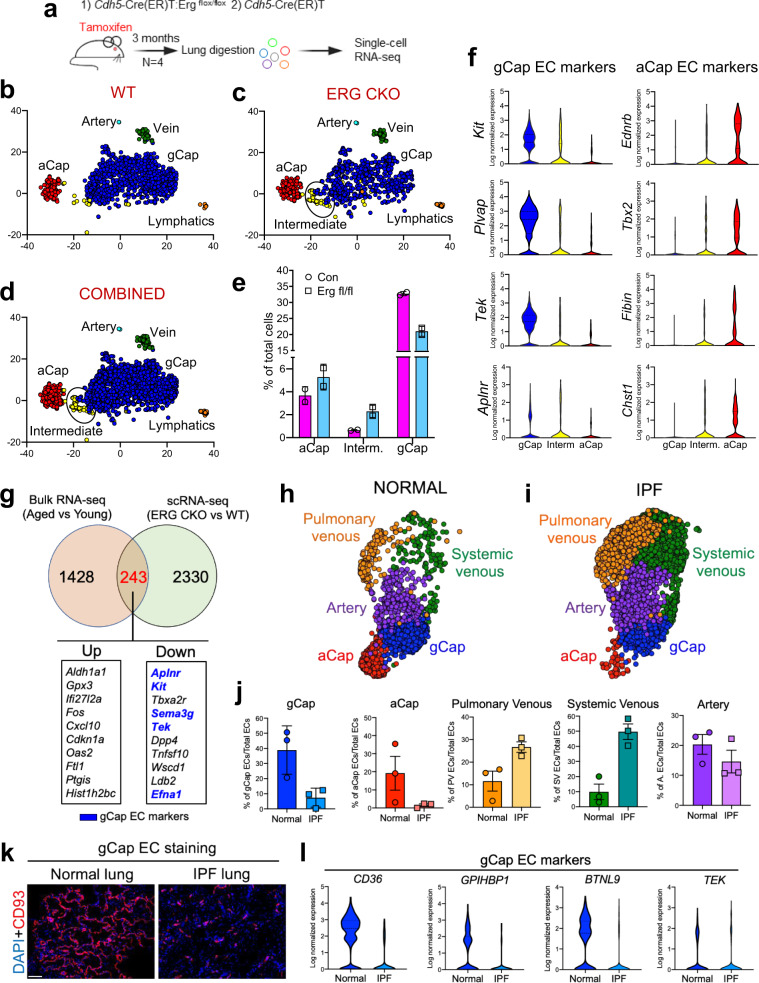
Vascular dysfunction is a hallmark of chronic diseases in elderly. The contribution of the vasculature to lung repair and fibrosis is not fully understood. Here, we performed an epigenetic and transcriptional analysis of lung endothelial cells (ECs) from young and aged mice during the resolution or progression of bleomycin-induced lung fibrosis. We identified the transcription factor ETS-related gene (ERG) as putative orchestrator of lung capillary homeostasis and repair, and whose function is dysregulated in aging. ERG dysregulation is associated with reduced chromatin accessibility and maladaptive transcriptional responses to injury. Loss of endothelial ERG enhances paracrine fibroblast activation in vitro, and impairs lung fibrosis resolution in young mice in vivo. scRNA-seq of ERG deficient mouse lungs reveales transcriptional and fibrogenic abnormalities resembling those associated with aging and human lung fibrosis, including reduced number of general capillary (gCap) ECs. Our findings demonstrate that lung endothelial chromatin remodeling deteriorates with aging leading to abnormal transcription, vascular dysrepair, and persistent fibrosis following injury.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
