

Integrin α3β1 promotes vessel formation of glioblastoma-associated endothelial cells through calcium-mediated macropinocytosis and lysosomal exocytosis
- PMID: 35879332
- PMCID: PMC9314429
- DOI: 10.1038/s41467-022-31981-2
Integrin α3β1 promotes vessel formation of glioblastoma-associated endothelial cells through calcium-mediated macropinocytosis and lysosomal exocytosis
Abstract
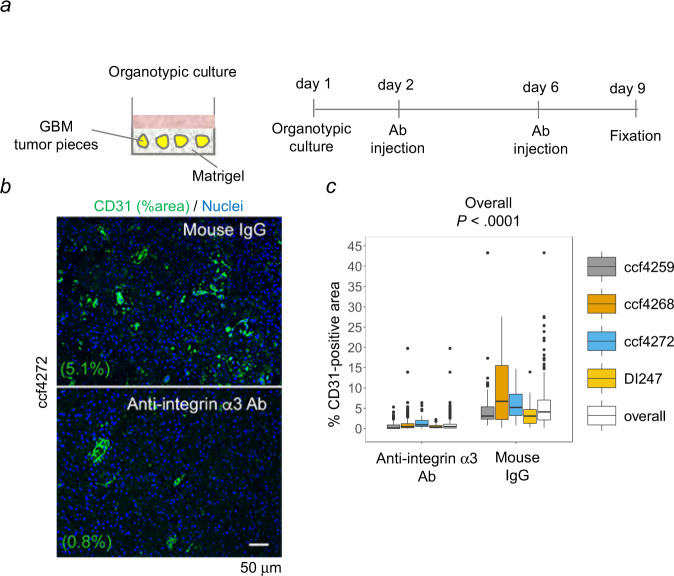
Therapeutic targeting of angiogenesis in glioblastoma has yielded mixed outcomes. Investigation of tumor-associated angiogenesis has focused on the factors that stimulate the sprouting, migration, and hyperproliferation of the endothelial cells. However, little is known regarding the processes underlying the formation of the tumor-associated vessels. To address this issue, we investigated vessel formation in CD31+ cells isolated from human glioblastoma tumors. The results indicate that overexpression of integrin α3β1 plays a central role in the promotion of tube formation in the tumor-associated endothelial cells in glioblastoma. Blocking α3β1 function reduced sprout and tube formation in the tumor-associated endothelial cells and vessel density in organotypic cultures of glioblastoma. The data further suggest a mechanistic model in which integrin α3β1-promoted calcium influx stimulates macropinocytosis and directed maturation of the macropinosomes in a manner that promotes lysosomal exocytosis during nascent lumen formation. Altogether, our data indicate that integrin α3β1 may be a therapeutic target on the glioblastoma vasculature.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
